


Abstract
A polymorphism involving a variable number of tandem repeats (VNTR) has been described in the 3′ untranslated region of the gene (SLC6A3) coding for the dopamine transporter (DAT). This polymorphism has 2 common alleles, designated as 10-repeat (∗10R) and 9-repeat (∗9R), that have been linked with several human clinical phenotypes. Previous investigations of the effects of the SLC6A3 polymorphism on DAT availability in smaller samples of humans have yielded divergent results. Methods: We assessed genotype at the SLC6A3 promoter VNTR polymorphism in 96 healthy European Americans (age range, 18–88 y) who also underwent SPECT with 123I-2β-carbomethoxy-3β-(4-iodophenyl)tropane (β-CIT) for measurement of striatal DAT protein availability. A ratio of specific to nondisplaceable brain uptake (i.e., V3″ = [striatal -occipital]/occipital), a measure proportional to the binding potential, was derived. For this analysis, 9–9 homozygotes and 9–10 heterozygotes were grouped as SLC6A3∗9R carriers and contrasted with SLC6A3∗10R homozygotes. Results: The SLC6A3∗9R carriers had significantly higher striatal DAT availability (V3″) than did the SLC6A3∗10R homozygotes, controlling for age (F1,93 = 6.25, P = 0.014, analysis of covariance). The ∗9R carriers (n = 41, 49.8 ± 19.5 y) had a mean increase in striatal DAT availability of 8.9% relative to the ∗10R homozygotes (n = 53, 49.9 ± 19.2 y). Striatal subregion analysis revealed that the effect of DAT genotype was significant for both the caudate and the putamen. Conclusion: These results support the interpretation of higher DAT levels in association with the ∗9R allele in European Americans and may relate to previously observed associations between DAT genotype and neuropsychiatric diseases.
A polymorphism involving a variable number of tandem repeats (VNTR) has been described in the 3′ untranslated region of the gene (SLC6A3) coding for the dopamine transporter (DAT). This polymorphism has 2 common alleles with either 9 (∗9R) or 10 (∗10R) repeats of a 40-base-pair sequence; less common alleles range from 3 to 11 repeats. Each common allele has been linked with several human clinical phenotypes, including neuropsychiatric diseases associated with dysregulation of dopamine transmission. The ∗10R allele has been associated with attention deficit hyperactivity disorder (ADHD) in several studies (1–4). The ∗9R allele has been associated with a history of cocaine-induced paranoia among cocaine-dependent European Americans (5), severity of alcohol withdrawal symptoms in some (6,7) but not all (8) studies, and a reduced risk of tobacco smoking (9,10). The ∗9R allele has been investigated for and found not to be associated with polysubstance abuse (11), cocaine dependence (5), delusional disorder (12), schizophrenia (13), Tourette’s syndrome (14,15), or alcoholism itself (6,15). In vitro studies (16–18) have suggested that the SLC6A3 VNTR polymorphism influences gene expression, but results have been conflicting (17,18) regarding the relative gene expression of the ∗9R and ∗10R alleles.
With the advent of functional brain imaging has come the possibility of studying the effects of genetic polymorphisms in living subjects. Three recent neuroimaging investigations have used SPECT and the radioligand 123I-2β-carbomethoxy-3β-(4-iodophenyl)tropane (β-CIT) to examine the effects of the SLC6A3 polymorphism on DAT protein availability in humans. Heinz et al. (19) studied 14 abstinent alcoholics and 11 control subjects and found that SLC6A3 9–10 heterozygotes had a mean 22% decrease in DAT binding in putamen, but not caudate, compared with SLC6A3∗10R homozygotes. Jacobsen et al. (20) studied 27 healthy control subjects and reported a mean 13.4% greater striatal DAT availability in the SLC6A3∗9R carriers than in the SLC6A3∗10R homozygotes. Martinez et al. (21) studied 29 patients with schizophrenia and 31 healthy controls and found no significant association between VNTR polymorphism and DAT density. Most recently, Lynch et al. (22) used a different SPECT ligand, 99Tc-TRODAT-1, to examine 100 patients with Parkinson’s disease and 66 healthy controls and observed no relationship between DAT genotype and DAT protein availability.
The discrepant results in previous studies may be attributable to several factors, including sample size, different outcome measures and image analysis techniques, and diagnostic and population heterogeneity. We therefore sought to examine a large group that was diagnostically and racially homogeneous. In the present study, the effect of SLC6A3 genotype on DAT protein availability was measured in 96 healthy European Americans.
MATERIALS AND METHODS
Sample
The study population consisted of 96 healthy European American volunteers (54 male and 42 female) who ranged in age from 18 to 88 y (mean ± SD, 50 ± 19 y). They underwent a clinical examination by a research psychiatrist to exclude any neurologic or psychiatric disease, alcohol abuse, or substance abuse. The smoking status of most subjects was not documented. Screening procedures included a physical and neurologic examination, electrocardiography, serum chemistries, thyroid function studies, complete blood cell count, urinalysis, and urine toxicology screen. Women of childbearing potential were required to test negatively for pregnancy (serum at screening; urine immediately before tracer injection). Subjects at least 55 y old (n = 31) were also required to have no significant evidence of cognitive impairment, as indicated by a Mini-Mental State Examination (23) score of 27 or greater. Subjects at least 68 y old (n = 25) were required to have normal brain MRI findings. No subject was taking medication known to affect the brain dopamine system. All subjects gave written informed consent to the research protocol, which was approved by the Yale Human Investigation Committee and conducted in accordance with the Declaration of Helsinki.
To maximize the statistical power of the present healthy population, we included all subjects for whom 123I-β-CIT SPECT scans had been analyzed by a single rater using a uniform methodology and for whom DNA was available. This sample therefore contained subjects in common with several of our previous reports. It was a subset of the sample (n = 126) for which we previously reported an age-related decline in dopamine and serotonin transporters (24,25) using the same 123I-β-CIT SPECT scans. It also overlapped with the 2 earlier 123I-β-CIT studies by our group reporting genetic results. Specifically, it contained 12 subjects’ scans in common with Jacobsen et al. (20) and 17 subjects in common with Martinez et al. (21). All scans, including these common scans, were analyzed by a rater different from those in the previous 2 reports using a somewhat different methodology, as will be discussed.
DNA Analyses
Genotyping was accomplished as described previously (20). The number of repeats contained in each allele, determined from polymerase chain reaction product size, is reflected in the numeric designation of each allele.
SPECT Imaging
All subjects received 0.6 g of a saturated solution of potassium iodide in the 24 h before undergoing SPECT. They then received an injection of 123I-β-CIT (222 ± 26 MBq; specific activity > 185 million MBq/mmol) on day 1, followed 22.9 ± 1.9 h later by a 24-min scan with a Picker PRISM 3000 (n = 72) or 3000XP (n = 24) SPECT camera equipped with a low-energy, high-resolution fanbeam collimator (128 × 128 matrix, 120° angular range, 3° angular step, 40 steps, 36 s per step, 15.5-cm radius of rotation). In this configuration, the PRISM 3000 acquires images at a reconstructed resolution of 12.3 mm in full width at half maximum as determined by an 123I point source in water. Comparability of the 2 cameras was confirmed by imaging an anthropomorphic striatal phantom on both instruments (using identical filtration, reconstruction, attenuation correction, and regions of interest [ROIs] and demonstrating 2%–4% variability between instruments in the striatal-to-background ratio), as well as by imaging 26 subjects on both cameras from a single 123I-β-CIT injection as described previously (25). Previous studies have demonstrated that 123I-β-CIT reaches equilibrium binding in the brain by 18–24 h (26), yielding a simple unitless ratio of regional radioactivities (V3″ = specific/nondisplaceable binding = [striatal -occipital]/occipital) in estimating DAT availability. Before scanning began, 4 or 5 fiducial markers filled with 185 kBq of Na99mTcO4 were attached to the skin along the canthomeatal plane to identify this plane during image analysis.
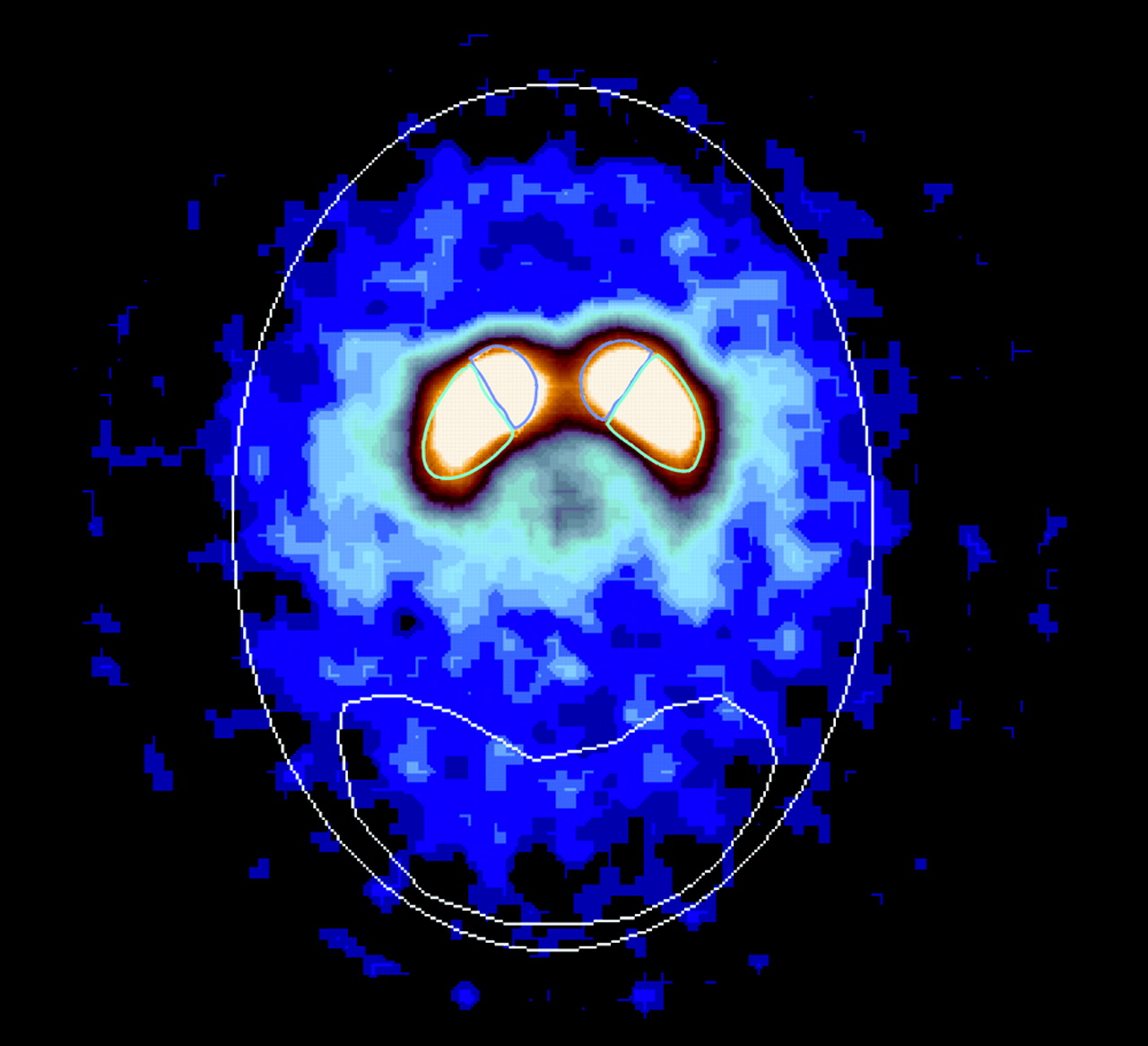
Images were reconstructed from photopeak counts (159 ± 16 keV) using standard filtered backprojection (Butterworth; power, 10; cutoff, 1.16 cm−1) and displayed as a 128 × 128 × 64 matrix with a voxel size of 2.07 × 2.07 × 3.56 mm (15.25 mm3). Subsequent image analysis was performed by an operator who was unaware of subject demographics. SPECT data were reoriented to correct for deviations from the canthomeatal plane, as identified by the fiducial markers. Eight contiguous transaxial slices with the highest uptakes in striatum were identified from a reconstructed midsagittal image and digitally summed to yield a transaxial slice 28.5 mm thick. Attenuation correction was performed using a Chang zero-order method (attenuation coefficient, μ, = 0.15 cm−1) within an ellipse drawn around the skull. Standard ROI templates for left and right caudate (424 voxels or 6.5 mL each), left and right putamen (824 voxels or 12.6 mL each), and occipital cortex (7,912 voxels or 120.6 mL) were positioned on the summed slice (Fig. 1). Correction for scatter or partial-volume effects was not attempted. Using similar methods with a different SPECT scanner, we have demonstrated excellent test-retest reproducibility in striatal V3″ for 123I-β-CIT (mean variability = 6.8% ± 6.8%; intraclass correlation coefficient = 0.96) (27).

Transaxial 123I-β-CIT SPECT image at level of striatum. Scan demonstrates ROIs positioned over right and left caudate, right and left putamen, and occipital cortex. Representative attenuation ellipse is also displayed.
Statistical Analysis
V3″ for striatum and striatal subregions (caudate and putamen) was computed without conversion of SPECT counts per minute to absolute units of radioactivity as ([cpm/voxel]striatal - [cpm/voxel]occipital)/(cpm/voxel)occipital. Given the relative rarity of 9–9 homozygotes, 9–9 homozygotes and 9–10 heterozygotes were grouped as SLC6A3∗9R carriers and contrasted with SLC6A3∗10R homozygotes, as has been done in previous studies (20,21). Statistical analyses were conducted using analysis of covariance (ANCOVA), controlling for age, which has been strongly correlated with striatal V3″ in previous studies with 123I-β-CIT (25,28). The proportion of variance explained by SLC6A3 genotype and age in the model was calculated as η2. The effect sizes of genotype and age in the model were calculated according to the formula B/(SEB , where B is the regression coefficient of the variable, SEB is the SE of B, n is the sample size, and 3 is the number of parameters in the model. To establish the independence of the present results from those of a previous positive report by our group (20), we performed a separate ANCOVA after removing 12 subjects whose scan data also were analyzed by a different method in that report. All statistical analyses used the SPSS (SPSS Inc.) or SYSTAT (SYSTAT Inc.) software packages and 2-tailed tests of significance.
, where B is the regression coefficient of the variable, SEB is the SE of B, n is the sample size, and 3 is the number of parameters in the model. To establish the independence of the present results from those of a previous positive report by our group (20), we performed a separate ANCOVA after removing 12 subjects whose scan data also were analyzed by a different method in that report. All statistical analyses used the SPSS (SPSS Inc.) or SYSTAT (SYSTAT Inc.) software packages and 2-tailed tests of significance.
RESULTS
Genotype Composition of Sample
Two subjects with rare genotypes (7–10 and 10–11) were excluded from subsequent analyses. Frequencies of VNTR genotypes for the remaining 94 subjects are displayed in Table 1. Of these, 41 were SLC6A3∗9R carriers (36 were 9–10 and 5 were 9–9) and 53 were SLC6A3∗10R homozygotes. Overall frequencies of the ∗9R and ∗10R alleles, 24% and 75%, respectively (for all 96 subjects), were similar to those observed by Vandenbergh et al. (29) (24% and 70%, respectively) and those observed in homogeneous European Americans by Doucette-Stamm et al. (30) (27% and 72%, respectively) and Gelernter et al. (31) (28% and 71%, respectively). The distribution of the 3 common genotypes did not differ significantly from Hardy-Weinberg equilibrium expectations (Table 1).
Frequencies of VNTR Polymorphism of DAT Gene (SLC6A3) in Healthy European Americans
Effect of Genotype on DAT Availability in Striatum
Striatal 123I-β-CIT SPECT data are displayed in Figure 2. The SLC6A3∗9R carriers had significantly higher striatal DAT availability (V3″) than did the SLC6A3∗10R homozygotes, controlling for age (F1,93 = 6.25, P = 0.014, ANCOVA), and the effect of age was significant in the ANCOVA model (t93 = −8.80, P < 0.001). The addition of sex as a covariate would not alter the results (F1,93 = 6.21, P = 0.015), nor would the effect of sex be significant in the ANCOVA model (t = 0.80, P = 0.43). The ∗9R carriers (n = 41, 49.8 ± 19.5 y) had a mean increase in striatal DAT availability of 8.9% relative to the ∗10R homozygotes (n = 53, 49.9 ± 19.2 y). The proportion of variance (η2) explained by SLC6A3 genotype and age in the model was calculated as η2SLC6A3 = 0.064 and η2age = 0.46. The effect sizes of genotype and age in the model were calculated as ESSLC6A3 = 0.26 and ESage = 0.92. Because the present findings replicated those of our earlier preliminary report (20), a separate analysis was conducted to establish the independence of the results. When the 12 subjects common to the 2 studies were removed, the effect of genotype remained significant in the ANCOVA model (F1,81 = 4.72, P = 0.033).
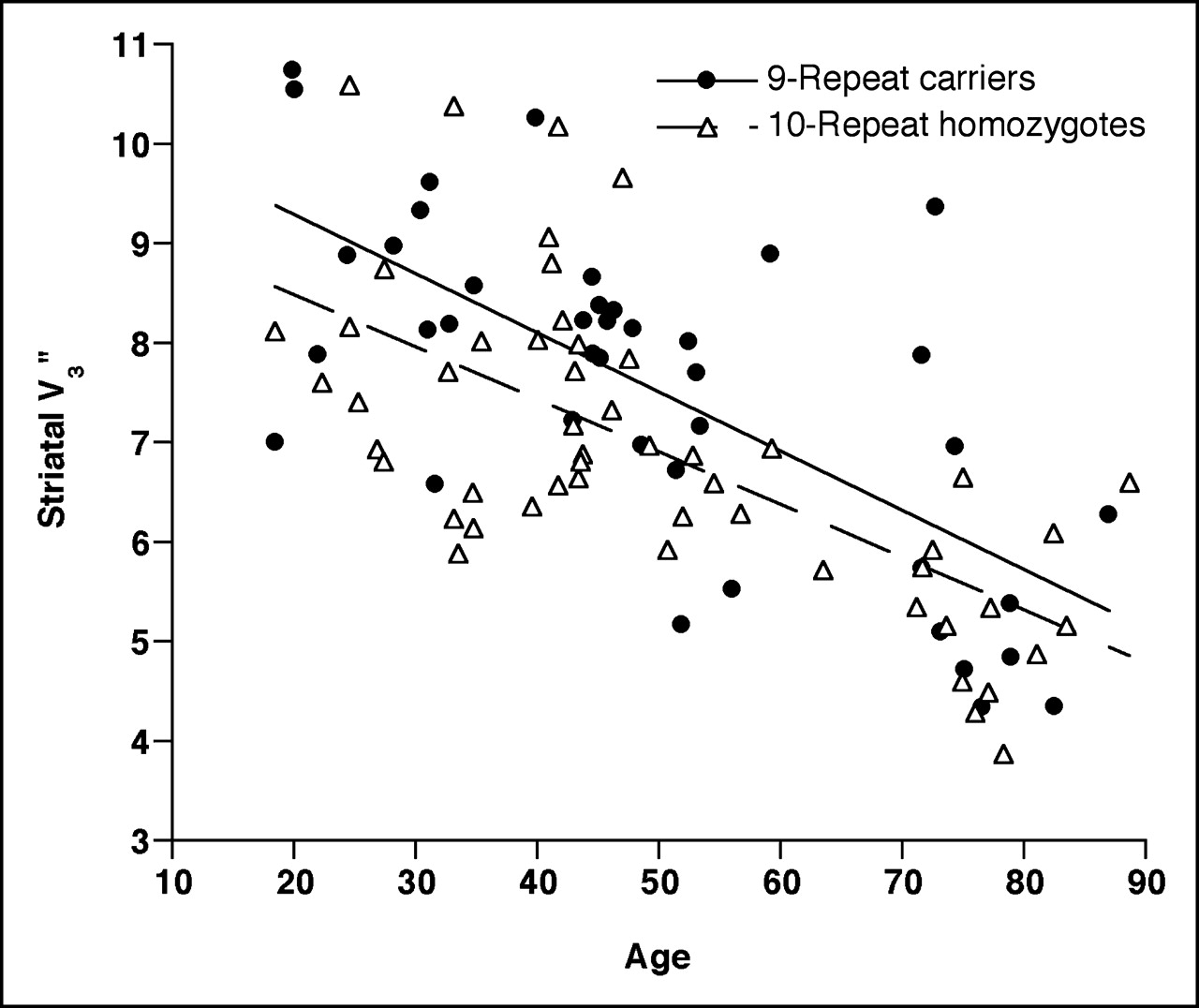
Striatal DAT availability (V3″) vs. age (in years) as measured by 123I-β-CIT SPECT in 94 healthy European Americans grouped according to SLC6A3 VNTR genotype. SLC6A3∗9R carriers (n = 41) had significantly higher striatal DAT availability (V3″) than did SLC6A3∗10R homozygotes (n = 53), controlling for age (F1,93 = 6.25, P = 0.014, ANCOVA). Both genotypic subgroups showed similar aging effects: For ∗9R carriers, V3″ declined by 44% over age range 18–88 y, or approximately 6.3% per decade. For ∗10R homozygotes, V3″ declined by 43% over age range 18–88 y, or approximately 6.2% per decade.
When the effect of genotype was instead considered as a 3-group ANCOVA, the main effect of genotype remained significant (F2,90 = 3.18, P = 0.046). Post hoc Tukey pairwise comparisons revealed a significant difference between the 9–10 and 10–10 groups (P = 0.037) but not between the 9–9 and 9–10 groups (P = 0.91) or the 9–9 and 10–10 groups (P = 0.74). However, this analysis was limited by the presence of only 5 subjects in the 9–9 group.
Effect of Genotype on DAT Availability in Striatal Subregions
When striatal subregions were examined separately (Fig. 3), the effect of genotype was again significant for both the caudate (F1,93 = 4.88, P = 0.030, ANCOVA; η2SLC6A3 = 0.051, η2age = 0.48; ESSLC6A3 = 0.23, ESage = 0.96) and the putamen (F1,93 = 6.92, P = 0.010; η2SLC6A3 = 0.071, η2age = 0.45; ESSLC6A3 = 0.28, ESage = 0.90). The ∗9R carriers had a mean increase in DAT availability of 7.8% for caudate and 9.5% for putamen relative to the ∗10R homozygotes.
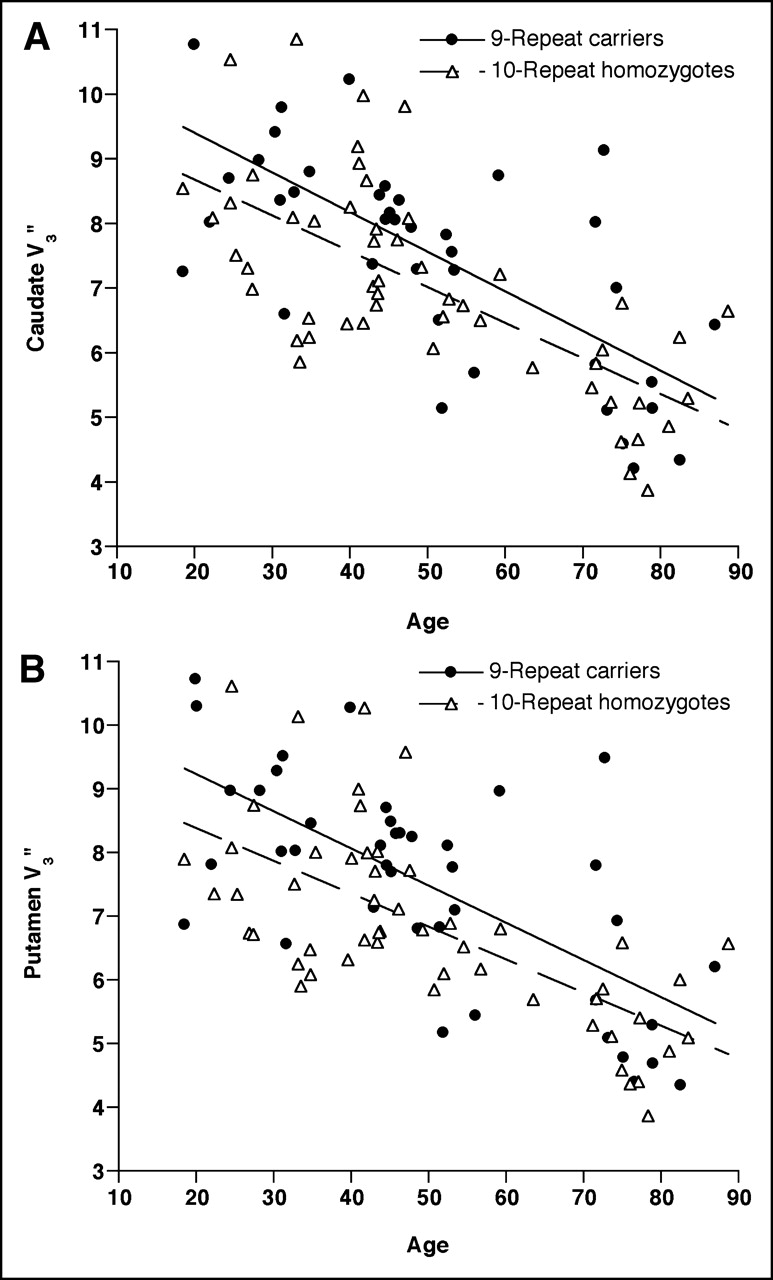
DAT availability (V3″) in caudate (A) and putamen (B) vs. age (in years) as measured by 123I-β-CIT SPECT in 94 healthy European Americans grouped according to SLC6A3 VNTR genotype. SLC6A3∗9R carriers (n = 41) had significantly higher DAT availability (V3″) in both caudate (F1,93 = 4.88, P = 0.030, ANCOVA) and putamen (F1,93 = 6.92, P = 0.010, ANCOVA), controlling for age.
Comparisons between left and right striatal regions showed significantly higher values for DAT availability (V3″) in the left (7.61 ± 1.88) than in the right (7.49 ± 1.82) putamen for both ∗9R carriers (left, 7.59 ± 1.68; right, 7.40 ± 1.70; t40 = 3.54, P = 0.001, paired t test) and ∗10R homozygotes (left, 6.90 ± 1.53; right, 6.78 ± 1.47; t52 = 2.80, P = 0.007). However, differences between the left and right caudate only approached statistical significance for both ∗9R carriers (left, 7.63 ± 1.72; right, 7.52 ± 1.70; t40 = 1.58, P = 0.12) and ∗10R homozygotes (left, 7.08 ± 1.57; right, 6.98 ± 1.56; t52 = 1.91, P = 0.06).
Aging Effects by Genotype
The effect of age on DAT availability, well documented in previous studies (25,28,32), was similar for the 2 genotypic subgroups (Fig. 2). Among the SLC6A3∗9R carriers, there was an age-dependent decline in striatal V3″ values (r39 = −0.69, P < 0.0001, V3″ = −0.059 × age + 10.48) of 44% over the age range 18–88 y, or approximately 6.3% per decade. Among the SLC6A3∗10R homozygotes, a similar decline of 43% (r51 = −0.67, P < 0.0001, V3″ = −0.053 × age + 9.55) was observed from age 18–88 y, or approximately 6.2% per decade.
DISCUSSION
We investigated the effect of SLC6A3 genotype on DAT availability in a large sample of diagnostically and racially homogeneous subjects: 96 healthy European Americans. When the contribution of age was controlled for, the SLC6A3∗9R carriers had significantly higher striatal DAT availability than did the SLC6A3∗10R homozygotes. This study provides a replication in an independent sample of the previous preliminary report from our group (20). When the 12 subjects common to the 2 studies were removed, the effect of genotype remained significant in the ANCOVA model (F1,81 = 4.72, P = 0.033, ANCOVA). These studies were also independent from a methodologic standpoint, as they used different SPECT image analysts as well as different methods of SPECT image analysis.
These results differed from those of 3 previous studies, which found either a mean 22% lower DAT binding in putamen in SLC6A3 9–10 heterozygotes than in SLC6A3∗10R homozygotes (19) or no effect of SLC6A3 genotype on striatal DAT availability (21,22). The fact that Heinz et al. (19) found the opposite—that is, greater DAT binding in ∗10R homozygotes—in putamen but not caudate raised the possibility of regional variation in DAT expression within striatum. We therefore separately analyzed striatal subregions and still found significantly greater DAT availability for ∗9R carriers in both caudate (F1,93 = 4.88, P = 0.030) and putamen (F1,93 = 6.92, P = 0.010). The previously described asymmetry (left > right) of DAT availability in putamen (25,32,33) was found to be equally true for both genotypic subgroups.
The divergent results among these 5 studies are potentially attributable to several methodologic differences. Heinz et al. (19) used the outcome measure BP′ (a ratio of the specific tracer uptake to the free plasma 123I-β-CIT concentration at equilibrium), whereas the other studies used either V3″ (20,21) (a ratio of specific to nondisplaceable brain uptake at equilibrium) or a closely related specific uptake value (22). V3″ has the theoretic disadvantage of assuming equivalent nondisplaceable tracer uptake between subjects (or at least study groups) but may be more reproducible because it does not depend on plasma measurements. Our results are unlikely to be confounded by a systematic difference in nondisplaceable 123I-β-CIT uptake between ∗9R carriers and ∗10R homozygotes, although this possibility cannot be excluded. Heinz et al. (19) and Jacobsen et al. (20) used MRI coregistration for ROI placement, enabling the use of cerebellum as the reference region, whereas Martinez et al. (21) and we in the present study used occipital cortex as the reference region and Lynch et al. (22) used whole supratentorial brain as the reference region. Although all reference regions have relatively low DAT concentrations, cerebellum may have the advantage of less 123I-β-CIT binding to serotonin transporters (34). However, this difference is again unlikely to produce systematic differences between ∗9R carriers and ∗10R homozygotes. Only Jacobsen et al. (20) performed transmission scanning for measured attenuation correction, whereas Martinez et al. (21), Lynch et al. (22), and we in the present study assumed uniform attenuation within an ellipse drawn around the skull, and Heinz et al. (19) performed no attenuation correction (personal communication, 2003). However, the improved accuracy in 123I-β-CIT quantitation by measured, compared with uniform, attenuation correction appears to be small (35) and, in any case, would be unlikely to introduce systematic biases between genotypic groups. None of these 4 studies corrected for partial-volume effects. Although unlikely, it is conceivable that the SLC6A3 polymorphism is associated with differences in brain morphology, including striatal shape and volume, that could systematically confound the calculation of striatal DAT availability.
Other important differences among these 5 studies include varying diagnostic and racial compositions of subject groups. Whereas the present study and that of Jacobsen et al. (20) included only healthy subjects, Heinz et al. (19) studied 14 abstinent alcoholic subjects and 11 control subjects; Martinez et al. (21), 29 patients with schizophrenia and 31 healthy controls; and Lynch et al. (22), 100 patients with Parkinson’s disease and 66 controls. SLC6A3 allele frequency is known to vary across population groups (30,31), although no significant effects on DAT availability have been established for race or the interaction of race∗ genotype. Lacking sufficient racial subsamples to perform a systematic analysis of racial effects, we opted to restrict our analysis to our largest population subgroup—European Americans. All 4 of the other studies (19–22) included subjects of non-European ancestry, although none of those studies likely possessed the statistical power to rigorously address the effect of race on DAT availability. Future studies using larger samples of different racial groups will be necessary to address this question.
The SLC6A3 VNTR polymorphism is in a noncoding region and is not associated with variations in the DAT protein sequence (29). Therefore, the present results suggest that the polymorphism affects either DAT gene expression—by altering transcriptional or translational efficiency or messenger RNA stability—or DAT protein function. Alternatively, the SLC6A3 VNTR may be in linkage disequilibrium with another polymorphism that modifies DAT availability. Furthermore, other polymorphisms, including those not yet characterized, may be even more influential on DAT phenotype than are polymorphisms for the DAT gene itself.
Several in vitro studies have suggested that the SLC6A3 VNTR polymorphism may influence gene expression. Michelhaugh et al. (16) transfected the SLC6A3∗9R VNTR allele into an immortalized dopaminergic cell line and into dopamine neurons in neonatal rat midbrain slices and observed enhanced transcription. However, other studies that compared the relative gene expression of the ∗9R and ∗10R alleles have yielded disparate results. Miller and Madras (18) found that vectors containing the ∗9R allele produced significantly higher levels of (firefly luciferase) reporter gene expression than did analogous vectors containing the ∗10R allele. By contrast, Fuke et al. (17), using a similar reporter system, found that luciferase expression was significantly higher in vectors containing the ∗10R than in those containing the 7-repeat (∗7R) or ∗9R alleles. Collectively, therefore, the in vitro studies to date suggest that the SLC6A3 VNTR polymorphism affects the expression of the DAT but are conflicting regarding the relative strength of the ∗9R and ∗10R alleles.
If the effects of the SLC6A3 VNTR polymorphism on DAT availability in healthy subjects can be generalized to various disease states—and that generalizability has not been established—then our results may have some degree of relevance for DAT function in neuropsychiatric disorders. Our results suggest that the mechanism of the association of this polymorphism with several disorders could be altered levels of central DAT protein, influencing concentrations of extracellular dopamine. However, our results appear to be at odds with at least the most obvious interpretation of some previous clinical associations of this polymorphism. For example, the ∗9R allele has been linked with some phenomena—cocaine-induced paranoia (5), severity of alcohol withdrawal symptoms (6,7), and a reduced risk of tobacco smoking (9,10)—thought to be associated with increased synaptic dopamine. These conditions thus would be more directly explained by decreased levels of DAT protein to clear dopamine from the synaptic space. However, this interpretation fails to take into account any other alterations in brain function consequent to genetic alteration in DAT function. Presumably, differences in function of a critical brain protein such as DAT that are present from the earliest stages of development would induce homeostatic or other compensatory responses.
Several studies (1–4) have suggested an association between the SLC6A3∗10R allele and ADHD. In conjunction with the present results, the results of those studies might indicate reduced DAT availability in patients with ADHD. In fact, neuroimaging investigations of this question have yielded conflicting results, including increases (36,37), decreases (38), or no difference (39) in striatal DAT binding in adult ADHD patients. Because the SLC6A3 polymorphism explains less than 4% of the overall variance in ADHD symptoms (4), it is unlikely to account for a significant generalized alteration in DAT availability in patients with ADHD. However, the relationship between DAT genotype and DAT protein availability may be altered in ADHD or, at least, in symptom subtypes of the disorder (e.g., accounting for the fact that the SLC6A3 polymorphism explains the relatively greater variance of hyperactive-impulsive than inattentive symptoms (4)). Clearly, additional imaging studies of ADHD incorporating genetic analyses will be necessary to clarify the relationship between DAT genotype and DAT protein availability in this disorder.
To be sure, the effect of the SLC6A3 polymorphism on striatal DAT availability appears to be small (ESSLC6A3 = 0.26; proportion of variance explained, 6.4%), particularly in comparison to that of age in the present sample (ESage = 0.92; proportion of variance explained, 46%) or Parkinson’s disease in another study by our group (40) (ESParkinson’s disease > 1.0). Thus, consideration of this polymorphism is unlikely to significantly enhance research on DAT levels in normal aging or Parkinson’s disease. However, studies of neuropsychiatric disorders involving modest DAT alterations may need to consider an association with this polymorphism as a potential confounding or explanatory factor.
CONCLUSION
These results support the interpretation of higher DAT levels in association with the ∗9R allele in European Americans and may relate to some of the previously observed associations between DAT genotype and neuropsychiatric diseases.
Acknowledgments
The authors thank Heide Klumpp, Michelle Early, Eileen Smith, Gary Wisniewski, Louis Amici, Kristina Estok, and Ann Marie Lacobelle for excellent technical assistance and Larry Muenz, PhD, for statistical advice. This research was supported by the American Federation for Aging Research, generous gifts from Rose and Philip Hoffer, and funds from the Department of Veterans Affairs (Research Enhancement Award in Depression), the National Institute of Mental Health (R43-MH48243, MH58620, and K02-MH01387), and the National Institute on Drug Abuse (R01-DA12849 and R01-DA12690).
Footnotes
Received Sep. 8, 2004; revision accepted Jan. 5, 2005.
For correspondence or reprints contact: Christopher H. van Dyck, MD, Departments of Psychiatry and Neurobiology, Yale University School of Medicine, One Church St., Suite 600, New Haven, CT 06510.
E-mail: christopher.vandyck{at}yale.edu
REFERENCES
In this issue





{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- ADHD: relation between cognitive characteristics and DAT1 / DRD4 dopamine polymorphisms
- Dopaminergic Role in Regulating Neurophysiological Markers of Sleep Homeostasis in Humans
- Gene x Abstinence Effects on Drug Cue Reactivity in Addiction: Multimodal Evidence
- Multilocus Genetic Composite Reflecting Dopamine Signaling Capacity Predicts Reward Circuitry Responsivity
- Genetically Determined Interaction between the Dopamine Transporter and the D2 Receptor on Prefronto-Striatal Activity and Volume in Humans
- Variation in dopamine genes influences responsivity of the human reward system
- Striatal Dopamine Transporter Availability Associated with Polymorphisms in the Dopamine Transporter Gene SLC6A3
- Gene gene interaction associated with neural reward sensitivity
- Additive Effects of Genetic Variation in Dopamine Regulating Genes on Working Memory Cortical Activity in Human Brain