


Abstract
The aim of this study was to investigate the effect of deep-brain stimulation of the subthalamic nucleus (STN) on regional cerebral blood flow (rCBF) throughout the entire brain volume in patients with Parkinson’s disease and to evaluate which of the brain areas showing an rCBF increase during STN stimulation related significantly to the improvement in motor function. Methods: Ten consecutive Parkinson’s disease patients (6 men, 4 women; mean age ± SD, 59 ± 8 y) with bilateral STN stimulators underwent 3 rCBF SPECT examinations at rest: the first preoperatively and the second and third postoperatively (follow-up, 4.8 ± 1.4 mo) with STN stimulators on and off, respectively. The motor unified Parkinson’s disease rating scale, the Hoehn and Yahr disability scale, and the Schwab and England activities-of-daily-living scale were used to evaluate the clinical state under each condition. Statistical parametric mapping was used to investigate rCBF during STN stimulation in comparison with rCBF preoperatively and with STN stimulators off. Also evaluated with statistical parametric mapping was the relationship between rCBF and individual motor scores used as covariates of interest. Results: STN stimulation significantly changed rCBF in the right pre–supplementary motor area (pre-SMA), anterior cingulate cortex, and dorsolateral prefrontal cortex and in the medial Brodmann’s area 8 (BA8) as defined in the atlas of Talairach and Tournoux (P < 0.05 corrected for multiple comparisons). The rCBF in these areas increased from the preoperative condition to the stimulators-on condition and decreased again after the stimulators were switched off. A significant correlation was detected between the improvement in motor scores and the rCBF increase only in the right pre-SMA and in the anterior cingulate motor area (P < 0.005, uncorrected). Conclusion: According to the topographic organization of the primate STN, our study shows that stimulation of the STN leads to rCBF increases in the motor (pre-SMA), associative, and limbic territories (anterior cingulate) in the frontal cortex. The significant correlation between motor improvement and rCBF increase in the pre-SMA and the anterior cingulate motor area reinforces the hypothesis that STN stimulation in parkinsonian patients can potentiate the cortical areas participating in higher-order aspects of motor control.
Idiopathic Parkinson’s disease (PD) is the second most common neurodegenerative disorder, with a reported incidence of 20 per 100,000 and a lifetime risk of 1 in 40 (1). Because PD is increasingly common with age, its prevalence is expected to triple over the next 50 y with the aging of the population. Current treatment is primarily based on dopamine replacement therapy using the dopamine precursor levodopa. However, in most patients, after producing an initially satisfactory response, levodopa fails to adequately control the tremor, rigidity, and other symptoms that characterize PD.
The failure of levodopa and dopaminergic therapy to relieve the symptoms of parkinsonian patients over the long term, and the improvement in stereotactic techniques, have recently brought about renewed interest in neurosurgical approaches to the treatment of the disease. As an alternative to surgery that creates lesions, a reversible surgical treatment that does not create lesions consists of implantation of electrodes in the target structure for chronic high-frequency deep-brain stimulation (DBS) (2).
The stimulation technique was applied to the subthalamic nucleus (STN) of animal models of parkinsonism (3,4) and then to patients with PD (5), dramatically improving all motor symptoms to the same level as the best levodopa response. The STN takes up a strategic position in the pathophysiology of PD because its main connections are inputs from motor areas of the cortex and the external segment of the globus pallidus pars and outputs to both output components of the basal ganglia–thalamocortical circuits, that is, the globus pallidus pars interna and the pars reticulata of the substantia nigra (6,7). According to current models of the function of the basal ganglia motor circuit (8–10) and the changes that occur in PD (11,12), the mechanism underlying the clinical effect of subthalamic DBS is associated with reduction of the excessive drive of the STN to the globus pallidus pars interna and pars reticulata of the substantia nigra (13). This reduction should decrease the tonic inhibitory influence of these structures on the activity of motor thalamic relay nuclei, resulting in activation of the motor cortical system and in a nearly normal state (14).
SPECT with perfusion tracers is widely available and may prove to be a sensitive diagnostic and research tool for investigating the discrete effects of STN stimulation on neural systems and the regionally specific correlates of clinical symptoms throughout the brain. Techniques used in SPECT imaging of regional cerebral blood flow (rCBF) have changed considerably over the past 10 y as more accurate and powerful means of measurement have become available. Advances have allowed robust and objective computation of parametric maps of regional activity in the human brain (15). These maps can be statistically analyzed at the voxel level, given reliable techniques for the spatial normalization of brain volumes in a standard stereotactic space and appropriate statistical tests (16,17). SPECT imaging and statistical parametric mapping (SPM) have been used to reveal changes in rCBF, as an index of synaptic activity, in patients with PD, deepening our understanding of the pathophysiologic correlates of parkinsonism (18).
Our purpose was to test the a priori hypothesis that, in accord with the current model of the basal ganglia motor circuit, the mechanism underlying the clinical effect of subthalamic DBS is associated with an rCBF increase in frontal motor areas and, particularly, in the supplementary motor cortex. Functional neuroimaging with perfusion SPECT and image standardization were used to address this issue. We first evaluated the effect that bilateral STN stimulation in PD patients had on rCBF throughout the entire brain volume. To this end, the SPM analysis was adapted for a multisubject analysis, and the pattern of brain areas with rCBF changes associated with STN stimulation was determined by comparing the stimulators-on condition with the preoperative condition and with the postoperative stimulators-off condition. We then evaluated which of the brain areas showing an rCBF increase when STN stimulators were on related significantly to the improvement in motor function.
This study was prospective, aiming to deepen our understanding of brain regions that may underlie the clinical benefits of DBS of the STN in parkinsonian patients. The results of this study also stressed the importance of imaging with SPECT as a suitable approach for making inferences about the neural correlates of brain disorders.
MATERIALS AND METHODS
Patients and DBS
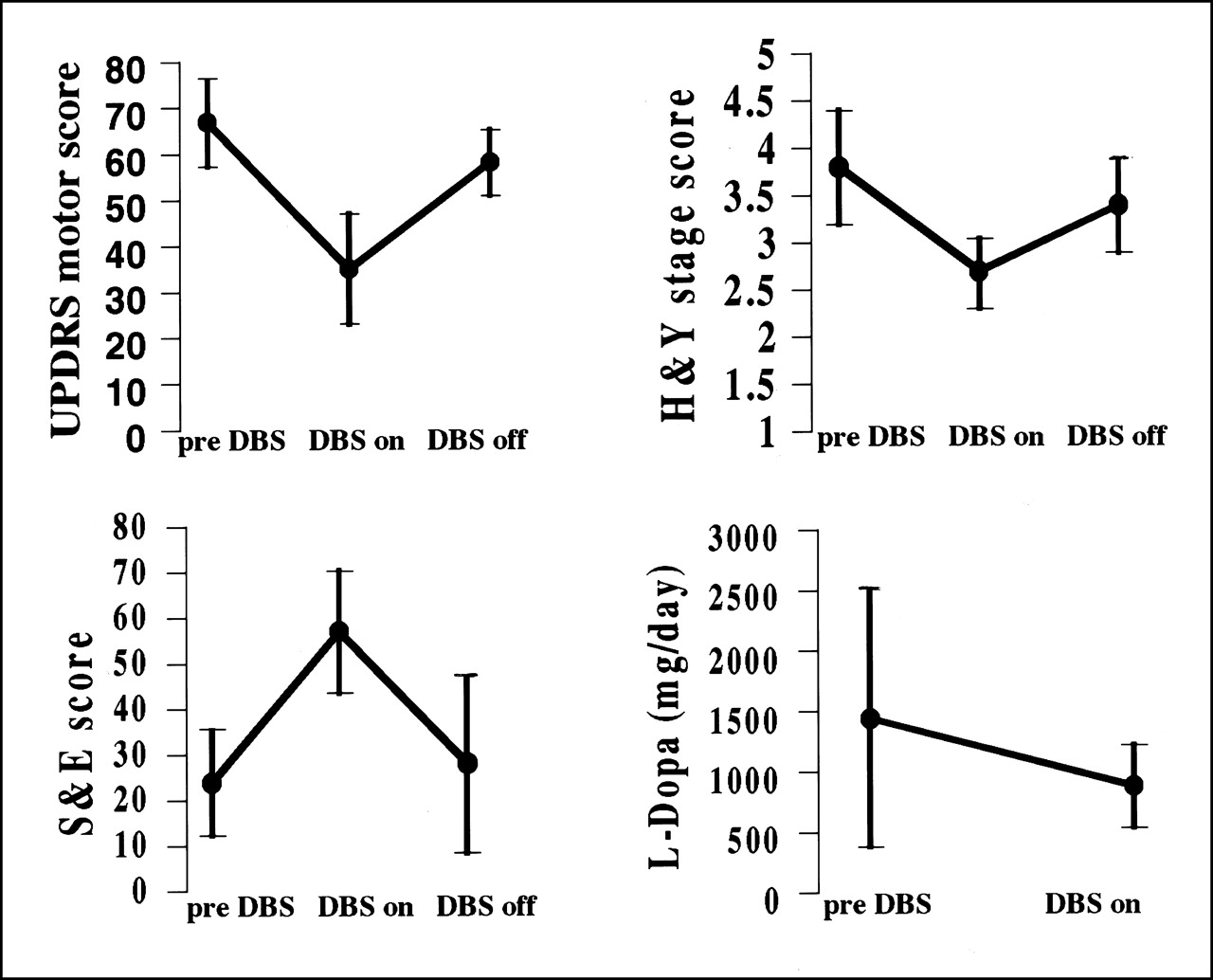
Ten consecutive patients with medically intractable PD, all levodopa responsive, operated on between 1998 and 2000 at our institute, were included in the study (4 women, 6 men; mean age ± SD, 59 ± 8 y; mean disease duration, 12 ± 4 y; range of disease duration, 6–18 y). The selection criteria for bilateral implantation of electrodes in the STN were the presence of clinically diagnosed idiopathic PD, disabling motor fluctuations despite all drug therapies, an age < 70 y, normal MRI findings for the brain, and no severe dementia (score on the Mini-Mental State Examination ≥ 24). The exclusion criteria were neurologic signs suggestive of secondary forms of parkinsonism and the presence of another significant medical illness. Patients were studied only after antiparkinsonian medications had been withdrawn for >12 h. Before scanning, and while unmedicated, patients were clinically evaluated with the motor unified Parkinson’s disease rating scale (UPDRS) (19), the Hoehn and Yahr disability scale (H&Y) (20), and the Schwab and England activities-of-daily-living scale (S&E) (21). At the time of surgery, the mean values for the motor UPDRS, H&Y, and S&E were 67 ± 9.6, 3.8 ± 0.6, and 24 ± 11.7, respectively (Fig. 1). The side prevalence of motor symptoms was left in 6 patients, bilateral in 3 patients, and right in 1 patient. All patients were treated with levodopa. The mean administered dose of levodopa before DBS was 1,445 ± 1,066 mg/d (Fig. 1). Four patients also received dopamine receptor agonist therapy (pergolide mesylate). The procedure for bilateral implantation of STN electrodes conformed with one previously published (22,23).

Plots of clinical scores (mean ± SD) for motor UPDRS, H&Y, and S&E for preoperative (before DBS) and postoperative conditions with STN stimulators on (DBS on) and off (DBS off) and plots of daily consumption of levodopa (L-Dopa) therapy before and after DBS implantation. Clinical scores were assessed in unmedicated patients. Reduction in motor UPDRS and H&Y scores and increase in S&E score indicate improvement in motor function, global stage disease, and performance of activities of daily living, respectively. Differences between mean scores for UPDRS, H&Y, S&E were significant (P < 0.001). Differences between mean scores of daily consumption of levodopa therapy were significant (P < 0.05). Results are expressed as mean ± SD
All patients gave written informed consent before implantation of the stimulation systems and SPECT scanning. The study was approved by the Ethics Committee of our institution.
Experimental Design
All patients underwent rCBF SPECT scanning 3 times at rest: once preoperatively, 1 d before surgery; once postoperatively with STN stimulators on, 4.8 ± 1.4 mo after surgery; and once 3 d later, with STN stimulators switched off 6 h before the scan and left off during the scan.
Data Acquisition
The patients received an intravenous dose of 99mTc-ethyl cysteinate dimer (bicisate, Neurolite; DuPont Merck Pharmaceutical Co., Billerica, MA) in a dimly lit room with minimal background noise. The administered dose was 740 MBq under each of the 3 scanning conditions. From 20 to 30 min after 99mTc-ethyl cysteinate dimer injection, SPECT data were acquired using a triple-head rotating gamma camera (PRISM 3000; Picker International Inc., Cleveland, OH) equipped with ultra-high-resolution fanbeam collimators. A polycarbonate head holder was used to reduce head movement during scanning. All patients were closely observed during scanning to detect head movement or dyskinesias. Acquisition consisted of 180 projections recorded in step-and-shoot mode over a 360° rotation arch (each head acquired 60 projections). The angular step was 2°, and the frame time was 22.50 s per step. Each acquisition was completed in 26 min. The acquisition matrix was 128 × 128, with a 3.5-mm sampling. All projection data were prefiltered with a bidimensional Butterworth filter for which cutoff frequency and order were adjusted for brain volume and counts of data projections. The currently applied values were a cutoff frequency of 0.28 cycle per pixel and an order of 15. The filtered data were reconstructed using an iterative algorithm based on conjugate gradients compensating for the spatial response of the collimator (24). This reconstruction method allowed for an in-plane and axial resolution of 5.5–5.7 mm in full width at half maximum. Attenuation correction with Chang’s method was performed for each slice, with a uniform attenuation coefficient of 0.11 (25). All reconstructed images were transferred to an AlphaStation workstation (Digital Equipment Corp., Maynard, MA) for further analysis.
Image Transformation and Statistical Analysis
All calculations and image transformations were performed using Analyze image display software (version 1.0; Mayo Clinic, Rochester, MN). Data were analyzed using SPM (SPM96; Wellcome Department of Cognitive Neurology, London, U.K.) implemented in MATLAB (The MathWorks, Inc., Natick, MA) (15). For each subject, the 3 scans were realigned with respect to the first baseline scan using the Automated Image Registration package (version 3.0; University of California, Los Angeles, CA) developed by Woods et al. (26). After realignment, all brain images were transformed into the standard stereotactic Talairach space (as described in the atlas of Talairach and Tournoux (27)). Spatial normalization involved a 12-parameter linear affine transformation and a nonlinear 3-dimensional deformation to match each scan to a provided reference image that already conformed to the standard space (16,28). As a result, each image was resampled into 2 × 2 × 2 mm with respect to the x-coordinate (right–left), y-coordinate (anterior–posterior), and z-coordinate (superior–inferior). These spatially normalized images were then smoothed with an isotropic gaussian kernel (full width at half maximum = 13 mm) to account for variations in gyral anatomy and individual variability in structure–function relationships and to increase the signal-to-noise ratio. Global blood flow was normalized by scaling across the entire dataset to a grand mean of 50 mL/100 mL per minute. The normalization process adjusted the rCBF values for each voxel to take into account variations in global activity across patients.
The pattern of brain areas with rCBF changes associated with STN stimulation was determined by comparing the adjusted rCBF voxel values when STN stimulators were on with the preoperative values and with the postoperative values when STN stimulators were off. Appropriate linear contrasts were used within a single paradigm to extract regions in which rCBF increased from the preoperative condition to the stimulators-on condition or decreased from the stimulators-on condition to the stimulators-off condition.
Moreover, covariance analysis was performed to extract regions whose changes in rCBF significantly correlated with changes in motor symptoms. To address this issue, individual motor scores during each scanning condition were introduced into the design paradigm. Global blood flow was used as a confounding covariate. These analyses generated SPM{t} maps that were subsequently transformed to SPM{Z} maps (16), and the level of significance of areas of rCBF change was assessed by the peak height of their foci using estimations based on the theory of gaussian fields (17). When the main effect of STN stimulation on the entire brain volume was investigated, significance was accepted if voxels survived a height threshold of P < 0.05 after correction for multiple comparisons. For covariance analysis, voxels within the brain regions hypothesized to modulate motor ratings during STN stimulation were considered significant at an uncorrected spatially restricted probability value of <0.01. According to the current model of basal ganglia motor circuits, the cortical areas hypothesized to be engaged with clinical scores during STN stimulation were the supplementary motor area (SMA), the motor area, and the premotor area. Subcortical areas hypothesized to be involved included the striatum, pallidum, and thalamus. The brain areas with rCBF changes were identified and listed according to stereotactic coordinates. The results were also displayed as 3 orthogonal projections of SPM{Z}.
Comparisons of UPDRS, H&Y, and S&E scores for each patient preoperatively and with STN stimulation on and off postoperatively were tested using ANOVA. Post hoc comparisons were performed with the Scheffé test. Differences between the mean scores of each group were considered significant at a level of P < 0.05.
RESULTS
Clinical Improvement and Medication
All motor ratings improved from the preoperative condition to the stimulators-on condition and worsened from the stimulators-on condition to the stimulators-off condition (Fig. 1). Mean improvement and worsening were 48% and 66%, respectively, for UDPRS (P < 0.001); 29% and 25%, respectively, for H&Y (P < 0.001); and 137% and 49%, respectively, for S&E (P < 0.001).
The mean administered dose of levodopa decreased by 40% from the preoperative to the postoperative condition (P < 0.05) (Fig. 1). A reduction in the daily consumption of dopaminergic medication (pergolide mesylate) was also observed.
rCBF
Main Effect of STN Stimulation.
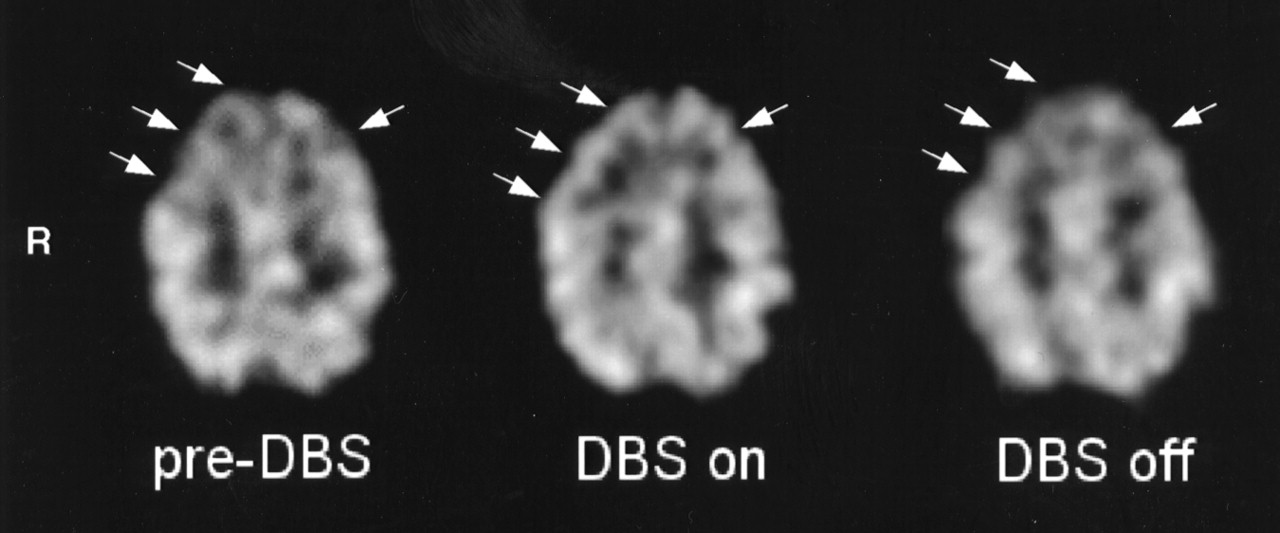
Significant rCBF changes were found in the medial part of the right superior frontal gyrus and in the upper part of the right medial frontal gyrus, corresponding to the rostral part of the SMA (right pre-SMA; medial BA6 in the atlas of Talairach and Tournoux (27)); bilaterally in the lower part of the medial frontal gyrus (BA8); in the anterior cingulate cortex (ACC) (BA32); and bilaterally in the lower part of the superior frontal gyrus, corresponding to the dorsolateral prefrontal (DLPFC)/frontopolar cortex (BA9/10). The region formed a continuous cluster containing 1,459 voxels with a peak of enhanced rCBF in the left BA10. The location and peak Z scores of the foci of rCBF increase are presented in Table 1. The extent of these activations is displayed in Figure 2A as SPM{Z} maximum-intensity-projection maps in 3 orthogonal views. The adjusted rCBF values of the voxels with the peak Z scores increased from the preoperative condition to the stimulators-on condition, and they decreased when the stimulators were switched off. The mean changes were 5.2% and 2.8%, 4.5% and 2.2%, 6.1% and 3.5%, and 5.5% and 2.2% for the pre-SMA (Fig. 2B), BA8, ACC, and DLPFC (Fig. 2C), respectively. No regions of significant rCBF decrease were evident. A representative SPECT study (patient 2) is displayed in Figure 3.
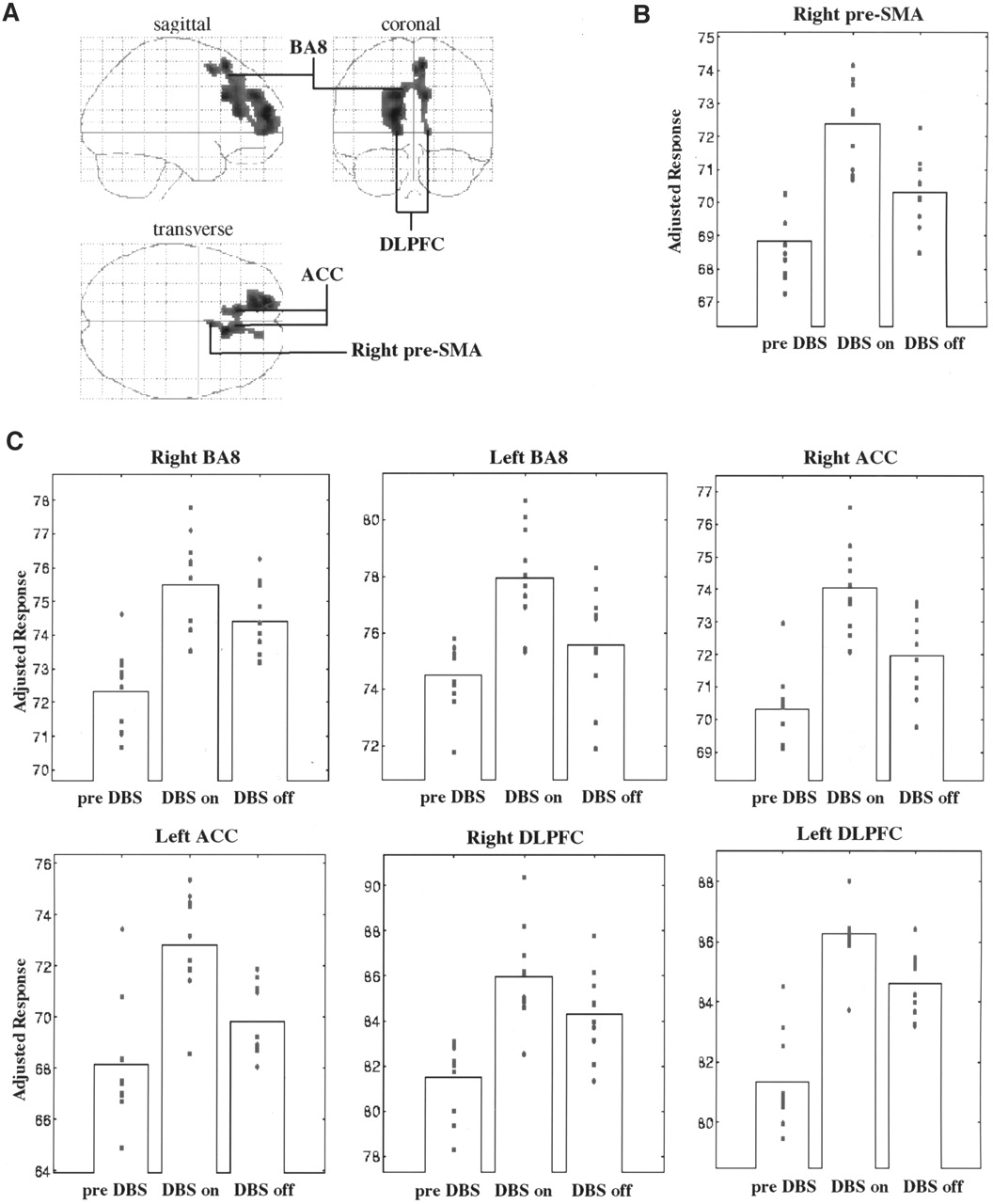
Depiction of cortical areas showing significant rCBF increase with STN stimulators on (DBS on) in comparison with preoperatively (pre-DBS) and postoperatively with STN stimulators off (DBS off). STN stimulation induced rCBF increases in motor (right pre-SMA), associative (DLPFC and medial BA8), and limbic (ACC) areas of frontal lobes in living brain of 10 parkinsonian patients. (A) Results are displayed as SPM projections in 3 orthogonal views, with cutoff of P < 0.05 corrected for multiple comparisons. (B and C) Plots of mean and individual adjusted rCBF values during preoperative (pre-DBS) and postoperative (DBS on and DBS off) conditions are depicted for right pre-SMA (B) and, bilaterally, for ACC, DLPFC, and medial BA8 (C).
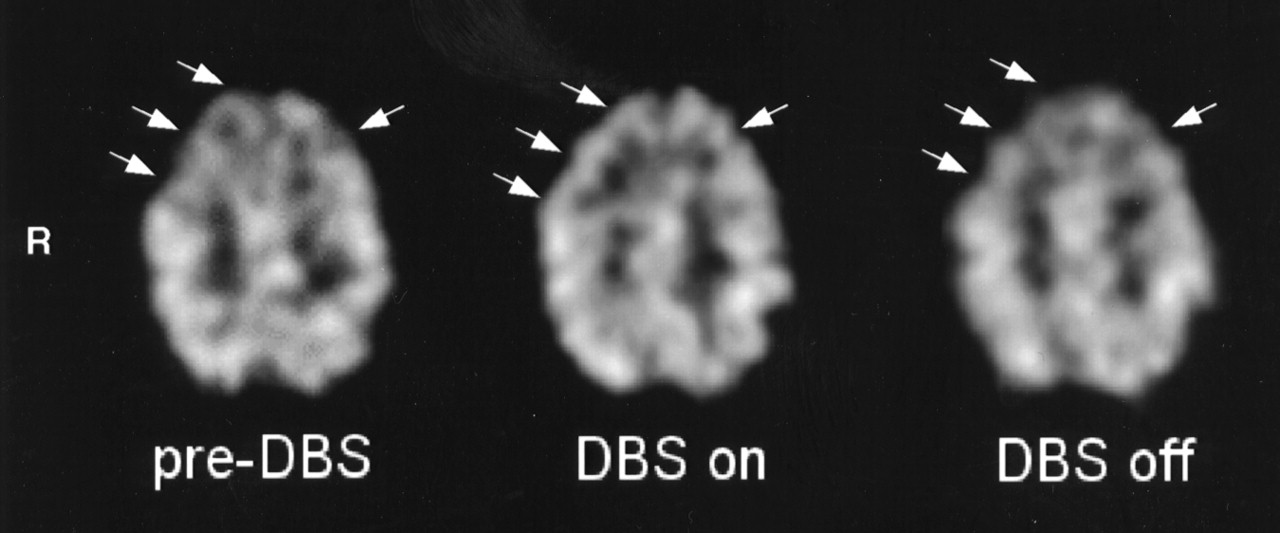
Transaxial brain SPECT section of patient 2 during each scanning condition: preoperatively (left), postoperatively with STN stimulators on (middle), and postoperatively with STN stimulators off (right). Arrows show rCBF increase in right frontal cortex and, to lesser extent, left frontal cortex and ACC during STN stimulation in comparison with preoperatively and during no STN stimulation.
Locations and Peak Z Scores of Areas with rCBF Increase During Bilateral DBS of STN (Main Effect of STN Stimulation)
Cerebral Perfusion Correlates of Motor Improvement.
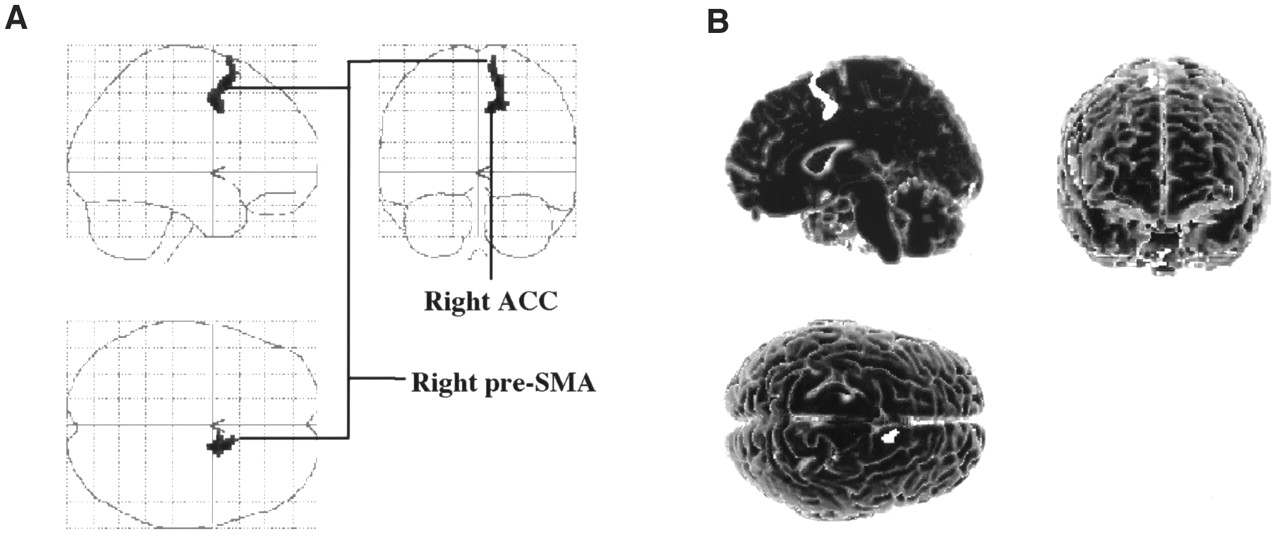
Covariance analysis showed a significant relationship (P < 0.005) between the rCBF and the motor scores in the right pre-SMA (medial BA6) and in an adjacent region of the right ACC (BA32). These regions formed a continuous cluster of 88 voxels with a peak of enhanced rCBF in the right medial BA6. These results are presented in Table 2 and Figure 4.
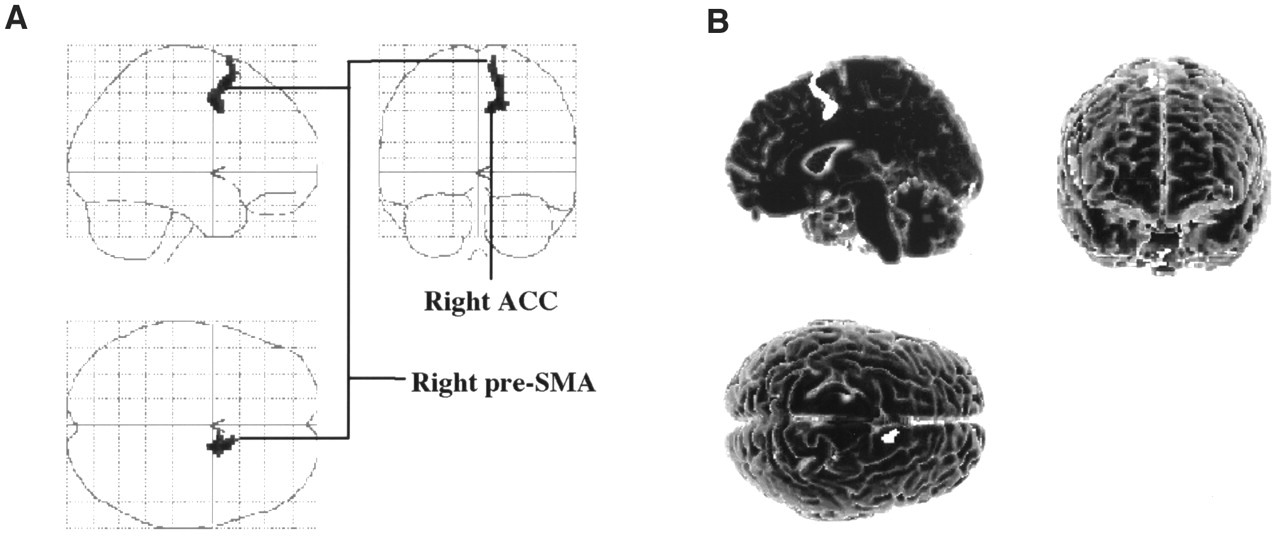
Cortical regions whose increases in rCBF significantly correlated with observed improvement in motor function during STN stimulation in comparison with preoperatively and during no STN stimulation. Significant relationship was detected between motor UPDRS scores and rCBF in right supplementary motor area (pre-SMA) and in small area of adjacent ACC referred to as anterior cingulate motor area. Results are displayed as SPM projections in 3 orthogonal views (A) and on 3-dimensional template (B), with cutoff of P < 0.005 (uncorrected).
Locations and Peak Z Scores of Areas with rCBF Increase During STN Stimulation That Significantly Correlated with Motor Improvement (Motor Scores)
DISCUSSION
In this study, we investigated rCBF changes in 10 PD patients who underwent clinically effective DBS of the STN. Motor ratings (UPDRS, H&Y, S&E) were significantly improved and levodopa assumption was significantly reduced in all 10 patients after DBS.
As far as rCBF changes are concerned, 2 main findings were observed. First, bilateral stimulation of the STN produced rCBF changes in several areas of the frontal lobes, including the pre-SMA, the ACC, the DLPFC, and the lower part of medial frontal gyrus, corresponding to the medial BA8 (Table 1; Figs. 2A and 3). The rCBF in these cortical regions increased from the preoperative condition to the stimulators-on condition and decreased after the stimulators were switched off, showing that the observed rCBF changes were related to the stimulation (Figs. 2B and 2C).
Second, when we investigated in which brain areas the rCBF changes correlated with motor ratings, significance was found for only a few of the above regions—specifically, for the pre-SMA and for a motor area of the adjacent ACC (Table 2; Fig. 4) (29,30).
Concerning the main effect of DBS stimulation, the finding that STN stimulation increases rCBF in the motor (pre-SMA), associative (DLPFC and medial BA8), and limbic (ACC) areas of the frontal cortex of the living human brain is consistent with the current view of both the topographic organization of the primate STN and basal ganglia–thalamocortical circuits. Indeed, based on STN neural connections, the motor, associative, and limbic territories have been identified in an orderly sequence from the dorsolateral to the ventrolateral portions of the STN (9). Moreover, the motor territory of the STN has been described to be somatotopically organized in the motor proper, SMA, and premotor territories (31). As first described by Alexander et al. (9), the topographic organization of the STN reflects the large-scale organization of the basal ganglia, a family of reentrant loops, each originating from a particular set of functionally related cortical fields in the frontal cortex (motor, associative, and limbic) and returning to parts of those same cortical fields by way of specific functionally and anatomically organized regions of the basal ganglia and thalamus. Accordingly, the outputs of the motor, associative, and limbic territories of the STN are directed to corresponding regions of the output stations of the basal ganglia, thalamus, and frontal cortex, providing important control of motor, cognitive, and emotional aspects of information processing (10).
Two previous 15O-H2O PET studies investigated the effect of monolateral STN stimulation on rCBF. The study of Limousin et al. (32) included 6 PD patients, all having a predominantly akinetic–rigid form of PD with severe motor fluctuations. In the study of Ceballos-Baumann et al. (33), the patient population consisted of 9 PD patients, all of whom initially experienced symptoms on 1 side, with that side being more affected than the other when stimulators were off. In both studies, the patients were scanned at rest and while moving a joystick during the stimulators-on and stimulators-off conditions. In the study of Limousin et al., when the effect of STN stimulation at rest was examined, stimulation that was effective produced significant rCBF changes in the sensorimotor (BA4) and lateral premotor (BA6) cortices. Similarly, Ceballos-Baumann et al. found rCBF changes in the primary motor cortex and in the globus pallidus–ventral thalamus associated with STN stimulation. These results agree with ours in that the SMA cortex of our SPECT study and the motor/premotor cortex of the other 2 PET studies strictly correspond to the frontal regions (9) in the basal ganglia motor circuit and were expected to be engaged during STN stimulation on the basis of the topographic organization of the motor territory of the STN (6).
The rCBF changes that we found in the motor, associative, and limbic cortical areas of the frontal lobes during STN stimulation were not unexpected. Indeed, the question of why DBS of the STN, which specifically targets the motor circuit, should affect other circuits, such as the associative and limbic, was already addressed by Jahanshahi et al. (34) in a recent review on the effect of DBS on executive function. According to their suggestions, the motor improvement observed in our study clearly showed that DBS was effectively targeting the motor part of STN, but the neural zone within which the functional effects of stimulation are active could be quite diffuse, extending across the motor, cognitive, and limbic territories of the STN. In addition, assuming that the effect of stimulation was confined to the motor part of the STN, another hypothesis is that rCBF changes in the motor, associative, and limbic cortical areas of the frontal lobes could result from interaction of the basal ganglia–thalamocortical circuits (35). Indeed, several studies have shown that the basal ganglia–thalamocortical circuits are functionally integrated and that interactions may occur at different levels of the circuits, including the STN.
When we investigated which of the brain areas showing an rCBF increase during the stimulators-on condition also showed a significant relationship with the observed improvement in motor function, we found the pre-SMA to show such a relationship. The rCBF increase also involved a small area of the posterior part of the ACC close to the pre-SMA, which has been referred to as the anterior cingulate motor area (29,30). The finding of rCBF increases in these cortical regions is consistent with our a priori hypothesis that STN stimulation may lead to a significant rCBF increase in the motor association cortex. In particular, this finding highlights that, although the main effect of subthalamic DBS on the frontal cortex was related to stimulation of all the territories—motor, associative, and limbic—expected to be engaged on the basis of the topographic organization of the STN (6,7), stimulation of the motor part of the STN seemed to be responsible for the dramatic improvement in motor function observed in our PD patients.
Our finding of a basal ganglia influence on the pre-SMA is of particular interest because this cortical region is associated with higher-order aspects of motor control, such as changing or updating plans for the future performance of motor behavior and motor imagery or learning (36). Abnormalities of these functions in PD, arising from excessive pallidal inhibition of SMA-related thalamic areas, has been reported to culminate in akinesia. As well as the pre-SMA, the motor anterior cingulate area has been reported to be involved in higher-order aspects of motor control, serving as a motivational motor interface between the limbic and motor systems (37). Our observation that STN stimulation can activate the pre-SMA and motor cingulate cortex and reverse parkinsonism is consistent with this view and reinforces the evidence that cardinal features of PD, such as akinesia, may be related mainly to underactivation of higher-order motor cortical areas. However, lower-order motor systems (i.e., red nucleus and cerebellar nuclei) are too small to be studied with this technique.
The increases in pre-SMA and cingulate cortex rCBF found in our study are consistent with the results of brain functional imaging studies performed on parkinsonian patients before and after treatment. First, the results of all these studies showed that before treatment, the pre-SMA and ACC were functionally underactive (37,38). Second, recovery of motor function, either by dopamine replacement therapy (38), by fetal nigral transplantation (40), or by pallidotomy (41), was associated with restoration of pre-SMA function. Our results confirm these findings also with DBS of STN, highlighting the crucial role of this area within the pathophysiology of PD.
In our study, SPM results were distinctly asymmetric when rCBF changes in motor cortical areas were observed (Figs. 2 and 4). Indeed, no rCBF changes were found in the left pre-SMA during STN stimulation. Interestingly, clinical symptoms were also asymmetric; left-sided symptomatology was present in 6 of 10 patients, and right-sided symptomatology was present in 1 patient. It is possible that the prevalence of left-sided motor symptoms might, in part, explain why rCBF increased in the contralateral pre-SMA when we switched on the STN stimulators. Nevertheless, it is also possible that the limited number of PD patients limited the ability of our study to reveal rCBF changes in the left pre-SMA.
CONCLUSION
In accord with the topographic organization suggested for the primate STN and the current model of basal ganglia–thalamocortical circuits, our study showed that stimulation of the STN leads to rCBF increases in the motor (pre-SMA), associative (DLPFC), and limbic (anterior cingulate) territories in the frontal cortex of PD patients. This study also showed a significant correlation between motor improvement and an rCBF increase in the pre-SMA bordering the anterior cingulate motor area during STN stimulation. In view of the functional connectivity of the basal ganglia–thalamocortical motor circuits and the results of 2 previous PET studies, this finding reinforces the hypothesis that, among nonprimary motor cortical areas, DBS of the STN in akinetic parkinsonian patients can potentiate the cortical areas participating in higher-order aspects of motor control. Our results also point out the utility of perfusion SPECT and SPM analysis for investigating the neural correlates of disease and for exploring the effects of therapy on the living human brain.
Acknowledgments
This study was partially funded by EC Contribution (contract PL 963130) and MURST 40%, 1999.
Footnotes
Received Sep. 17, 2001; revision accepted Jan. 31, 2002.
For correspondence or reprints contact: Alberto Pupi, MD, Nuclear Medicine Section, Careggi Hospital, University of Florence, Viale Morgagni 85, 50134 Florence, Italy.
E-mail: a.pupi{at}dfc.unifi.it





{kind=link}
{kind=link}
{kind=link}
{kind=link}