


Abstract
Monoclonal antibodies (mAbs) labeled with α-emitting radionuclides such as 211At, 212Bi, 213Bi, and 212Pb (which decays by β-emission to its α-emitting daughter, 212Bi) are being evaluated for their potential applications for cancer therapy. The fate of these radionuclides after cells are targeted with mAbs is important in terms of dosimetry and tumor detection. Methods: In this study, we attached various radionuclides that result in α-emissions to T101, a rapidly internalizing anti-CD5 mAb. We then evaluated the catabolism and cellular retention and compared them with those of 125I- and 111In-labeled T101. T101 was labeled with 211At, 125I, 205,6Bi, 111In, and 203Pb. CD5 antigen–positive cells, peripheral blood mononuclear cells (PBMNC), and MOLT-4 leukemia cells were used. The labeled T101 was incubated with the cells for 1 h at 4°C for surface labeling. Unbound activity was removed and 1 mL medium added. The cells were then incubated at 37°C for 0, 1, 2, 4, 8, and 24 h. The activity on the cell surface that internalized and the activity on the cell surface remaining in the supernatant were determined. The protein in the supernatant was further precipitated by methanol for determining protein-bound and non–protein-bound radioactivity. Sites of internal cellular localization of radioactivity were determined by Percoll gradient centrifugation. Results: All radiolabeled antibodies bound to the cells were internalized rapidly. After internalization, 205,6Bi, 203Pb, and 111In radiolabels were retained in the cell, with little decrease of cell-associated radioactivity. However, 211At and 125I were released from cells rapidly (211At < 125I) and most of the radioactivity in the supernatant was in a non–protein-bound form. Intracellular distribution of radioactivity revealed a transit of the radiolabel from the cell surface to the lysosome. The catabolism patterns of MOLT-4 cells and PBMNC were similar. Conclusion: 211At catabolism and release from cells were somewhat similar to that of 125I, whereas 205,6Bi and 203Pb showed prolonged cell retention similar to that of 111In. These catabolism differences may be important in the selection of α-radionuclides for radioimmunotherapy.
Recent radioimmunotherapy clinical trials using radiolabeled monoclonal antibodies (mAbs) directed against tumor-associated antigens have shown considerable promise. However, many uncertainties remain concerning the optimal implementation of this new modality. One of the most contentious issues is the selection of the optimal radionuclide. β-Emitters, such as 90Y and 131I, have been widely used (1–3).
Compared with β-emitters, α-emitters are more attractive for certain radioimmunotherapeutic applications. α-Emitters have several advantages over β-emitters, including higher linear energy transfer (which results in a relative biologic effectiveness of 5–20 times that of β-particles); reduced nonspecific irradiation to normal tissues around target cells because of their shorter pathlengths (60–100 μm); extremely high cytotoxicity as a result of DNA damage, which cells have a limited ability to repair; low dependence on dose rate and oxygen enhancement effects; and apoptotic mechanism resulting from α-radiation (4,5).
It has become increasingly evident that the behavior of radiolabeled catabolic products after total antibody degradation is an important factor for radioimmunotherapy. This fact was first recognized after comparison of antibodies labeled with radioiodine and 111In (6–8). Radioiodine, labeled to tyrosine residues of protein by standard iodination methods, is rapidly released from target cells after antibody internalization (9), suggesting that the protein is digested to amino acids and the iodine can be found as iodotyrosine or iodide on deiodination. In contrast, 111In-diethylenetriaminepentaacetic acid (DTPA) conjugated protein is retained within cells, probably within lysosomes (6,10) primarily as the lysine adduct, because of the cleavage of peptide bonds (11,12).
The fates of various radionuclides (111In, 125I, 90Y, 99mTc) attached to mAbs have been evaluated in in vitro cell systems after internalization (6,9,10). In some instances, these in vitro studies have been shown to be predictive of results in humans (7). In contrast, α-emitters have not been adequately assessed. In this study, we used the α-emitters 211At, 203Pb (surrogate for 212Pb that decays to 212Bi), and 205,6Bi (surrogate for 212Bi and 213Bi) to radiolabel mAb T101 and compared their internalization, metabolism, and retention with those of 125I- and 111In-labeled T101. 212Pb is the parent of 212Bi and has been used as an in vivo generator to deliver α-radiation through its daughter. Radioimmunoassay and Percoll gradient fractionation of cell organelles were used to test whether the retention of radioactivity was caused by lysosomal trapping of the radiometal-containing metabolites.
Materials and Methods
mAb and Radiolabeling
T101 is an IgG2a murine mAb specific for the CD5 antigen (13), which is a 65-kDa pan T-cell antigen expressed on normal T-lymphocytes and T-cell malignancies, with 30,000–180,000 sites per cell (14). This antibody is known to modulate rapidly once it binds to the CD5 on the cell surface, resulting in rapid internalization of the radiolabeled antibody. Modulation occurs in vivo and in vitro in malignant cell lines as well as in normal peripheral blood mononuclear cells (PBMNC) (15–17).
The T101 was purified from hybridoma ascites (Hybritech Inc., La Jolla, CA) and was radiolabeled with various radionuclides, including 211At, 205,6Bi (surrogate for 212Bi and 213Bi), and 203Pb (surrogate for 212Pb). For comparison with previous studies, T101 was also radiolabeled with 125I and 111In. 125I and 111In were obtained from New England Nuclear Life Science Products (Boston, MA). We directly radioiodinated T101 with 125I at a specific activity of 185–370 MBq/mg (5–10 mCi/mg) using the chloramine-T method (18). The nonbound 125I was removed by gel filtration chromatography. The radioactivity of these preparations was >95% protein bound, as determined by instant thin-layer chromatography or trichloroacetic acid (TCA) precipitation.
The T101 was conjugated with N-[(R)-2-amino-3-(p-isothiocyanatophenyl)propyl]-trans-(S,S)-cyclohexane-1,2-diamine-N,N,N′,N″,N‴-pentaacetic acid (CHX-A″-DTPA) and labeled with 111In or 205,6Bi at specific activities of 111–185 MBq/mg (3–5 mCi/mg) and 37–111 MBq/mg (1–3 mCi/mg), respectively, using methods previously described (19). Production and purification of 205,6Bi were performed at the National Institutes of Health (NIH) using a CS30 cyclotron with protons at 24–25 MeV (20). T101 was also conjugated with dodecanetetraacetic acid (DOTA) and labeled with 203Pb at specific activities of 74 MBq/mg (2 mCi/mg) (21,22). 203Pb-Chloride was obtained from Mount Sinai Medical Center (Miami, FL) and purified (19,23). For astatine labeling, N-succinimidyl N-(4-tributylstannylphenethyl) succinamate (SAPS) was first reacted with 211At and then purified and conjugated to the T101 antibody at specific activities of 37–111 MBq/mg (1–3 mCi/mg). The 211At was made at NIH using a 209Bi (α,2n) 211At reaction and separated and purified (24).
The stability of the various chelates for the radiometals used in vitro or in vivo has been described (22,25–27). Astatine-labeled antibodies using SAPS are stable in serum (unpublished data).
CD5 Positive Cells
In this study, normal PBMNC and MOLT-4, a human acute lymphocytic leukemia nonadherent tumor cell line, were used. The PBMNC were obtained from heparinized peripheral venous blood of healthy adults that was suspended in phosphate-buffered saline (1:2) and purified by Ficoll-Hypaque density gradient (LSM density gradient, Litton Bionetics, Kensington, MD) centrifugation. The resulting mononuclear cells were washed twice with phosphate-buffered saline (PBS) and resuspended in RPMI 1640 medium (Life Technologies Inc., Grand Island, NY) supplemented with 10% fetal calf serum (FCS; Life Technologies Inc.) (6). Typically, PBMNC resulting from these purifications have approximately 70% T-cells. MOLT-4 was also grown in RPMI 1640 medium with 10% FCS at 37°C in a moist atmosphere with 5% CO2 and cells were used while they were in log phase growth.
Immunoreactivity
The immunoreactivity of the various radiolabeled T101 preparations was determined using a single point modified cell binding assay at antigen excess (28). In brief, 5 ng radiolabeled T101 were incubated with 5,000,000 PBMNC cells for 2 h at 4°C and the cell-associated activity was separated and counted. Nonspecific binding was determined by adding 25 μg unlabeled T101.
Cell Binding, Internalization, and Catabolism of Radiolabeled Antibody
Target cells were washed in cold PBS and pelleted by centrifugation. The radiolabeled antibody was added to the cell pellets at a ratio of 1 μg antibody per 107 cells in a volume of 1 mL and incubated at 4°C for 1 h. After the 1-h incubation, the cells were washed twice with cold PBS to remove the unbound radioactivity. Aliquots containing 1 × 106 labeled cells were plated in 1 mL RPMI 1640 medium in microtiter plates, warmed to 37°C in a humidified 5% CO2 incubator for 0, 1, 2, 4, and 8 h (up to 24 h for the MOLT-4 cell line), and assayed for cell-associated radioactivity and supernatant radioactivity. All time points were done in duplicate or triplicate and all assays were repeated on at least 2 separate occasions, with the exception of 203Pb (because of its limited availability).
The internalized cell-associated fraction was determined with an acid wash method (29). Cell suspensions were centrifuged and the supernatant removed. After centrifugation, the cells were resuspended in 0.5 mL cold acid wash buffer (0.028 mol/L sodium acetate, 0.12 mol/L NaCl, 0.02 mol/L sodium barbital; pH 3.0). After a 6-min incubation period on ice, the cell suspension was centrifuged to separate the acid-soluble cell surface activity (supernatant) and the intracellular acid-resistant radioactivity (cell pellet) (29).
The extent of degradation of the radiolabeled mAb in the supernatant was evaluated. Culture supernatants (0.1 mL) were mixed with 0.5 mL methanol for 10 min to precipitate protein-bound radioactivity. After centrifugation, the radioactivity in the pellets (methanol precipitable) and supernatants (methanol soluble) was counted. Methanol precipitation was used rather than TCA precipitation because of reports of precipitation of astatide with the TCA, but not with methanol (30). In addition, methanol was preferred because Pb(II) in DOTA is also acid labile and could dissociate from the complex under acidic conditions.
Percoll gradient fractionation of organelles was performed. Cell aliquots were suspended in N-tris[hydroxymethyl]methyl-2-aminoethanesulfonic acid (TES) buffer (10 mmol/L triethanolamine, 0.25 mol/L sucrose; pH 7.5), disrupted with a Dounce homogenizer (monitored under microscopy), and sedimented for 10 min at 250g to remove nuclei and unbroken cells. The supernatant (0.8 mL) was layered on the surface of 20% solution of Percoll in TES buffer (7.2 mL) and centrifuged at 4°C for 60 min at 20,000g. Serial 0.5 mL fractions were collected from the top and assayed for radioactivity and for lysosomal galactosidase activity (10).
Statistical Analysis
Selected time points were compared using ANOVA and differences between 2 radionuclides were compared using the 2-tailed, unpaired t test.
Results
The immunoreactivity of the T101 radiolabeled with the various radionuclides showed high and specific bindability to target cells. Bindings to PBMNC are shown in Table 1. More than 70% of total added radioactivity bound to the cells and this binding was inhibited to low levels (<5%) with the addition of 25 μg unlabeled T101.
Immunoreactivity of 5 Radiolabeled T101s with PBMNC
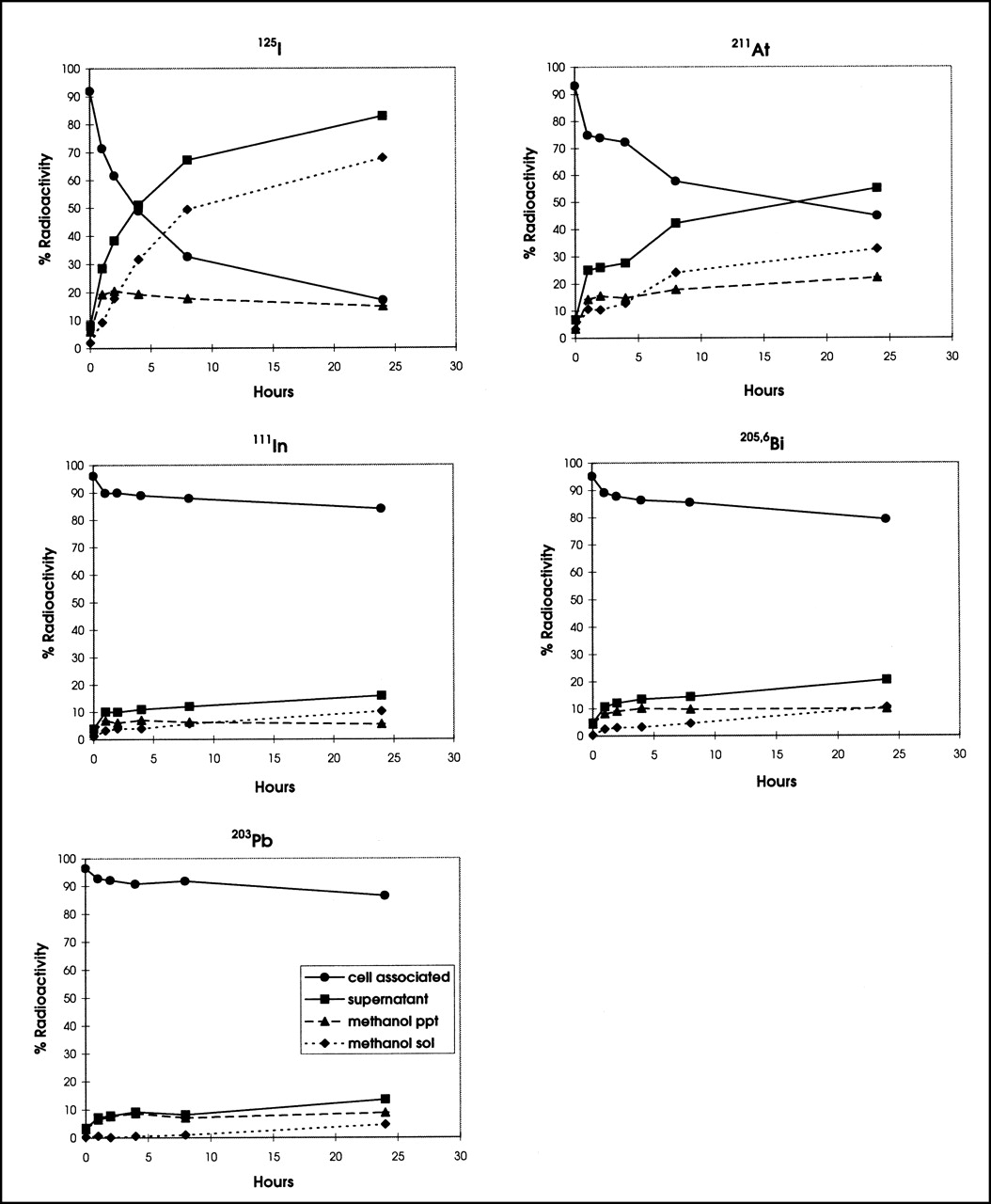
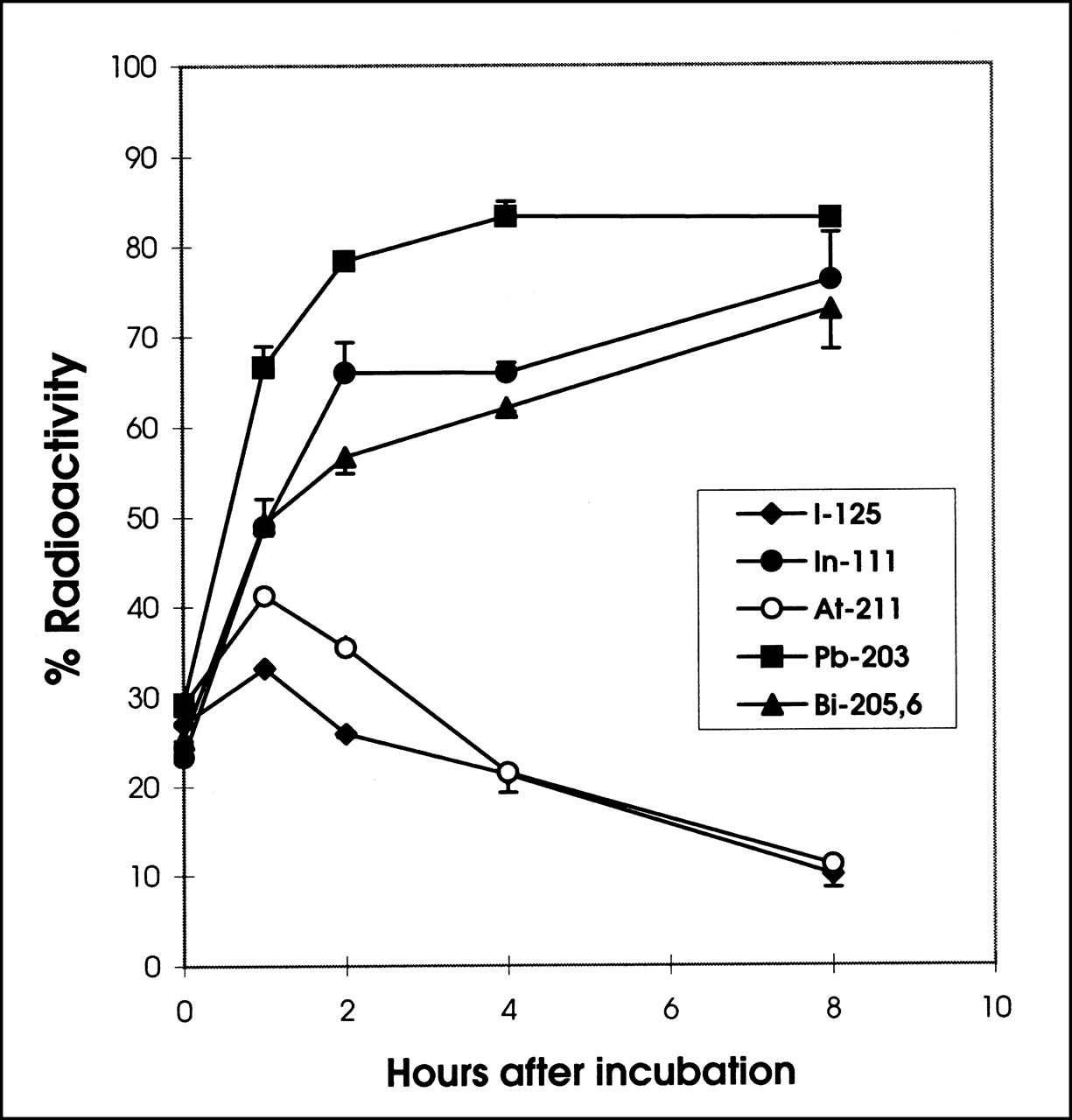
The catabolism of radiolabeled T101 antibodies by PBMNC and the fate of the radiolabels are shown in Figure 1. Large differences were seen in the cellular retention of the various radionuclides. As shown in Figure 1, the 125I cell-associated radioactivity decreased rapidly when incubated at 37°C, from >95% at the beginning to 87.58% ± 0.62%, 40.93% ± 0.81%, and 11.72% ± 0.61% of the original cell-bound 125I activity at 1, 4, and 8 h, respectively. The 125I in the supernatant increased accordingly and consisted mainly of non–methanol-precipitable 125I, consistent with small-molecular-weight products from a catabolized antibody. Although quantitatively different, 211At-T101 catabolism showed a pattern similar to that of 125I (Fig. 1), with rapid release from the cells: 74.28% ± 1.05%, 57.05% ± 0.75%, and 28.59% ± 0.83% remained cell-associated after incubation at 37°C for 1, 4, and 8 h, respectively. The majority of the released 211At was also in a non–protein-bound form. In contrast, the 111In-, 205,6Bi-, and 203Pb-radiolabeled T101 showed prolonged retention of these radiolabels by the PBMNC (Fig. 1). The cell-associated activity was 85.34% ± 0.69% and 89.10% ± 0.69% for 111In and 205,6Bi, respectively, after incubation for 8 h at 37°C. This was significantly higher than the retention of 125I or 211At (P < 0.0001 for 125I or 211At vs. either 111In or 205,6Bi). In the case of the metallic radionuclides, a grossly similar fraction of radioactivity was found in the protein-bound component as in the non–protein-bound fraction; however, for 125I- and 211At-T101, the amount in the non–protein-bound fraction was much higher in the supernatant.

Retention and release of 125I-, 211At-, 111In-, 205,6Bi-, and 203Pb-labeled T101 by PBMNC were determined. PBMNC (106) were incubated with 100 ng trace-labeled T101 at 4°C for 1 h and then washed, placed in fresh culture medium, and incubated at 37°C for 0, 1, 2, 4, and 8 h before quantifying amounts of cell-associated radioactivity and amount released into supernatant. Supernatant radioactivity was fractionated into methanol-precipitable noncatabolized component and methanol-soluble catabolized fraction. SD bars are plotted for replicates but are too small to be visualized for most time points.
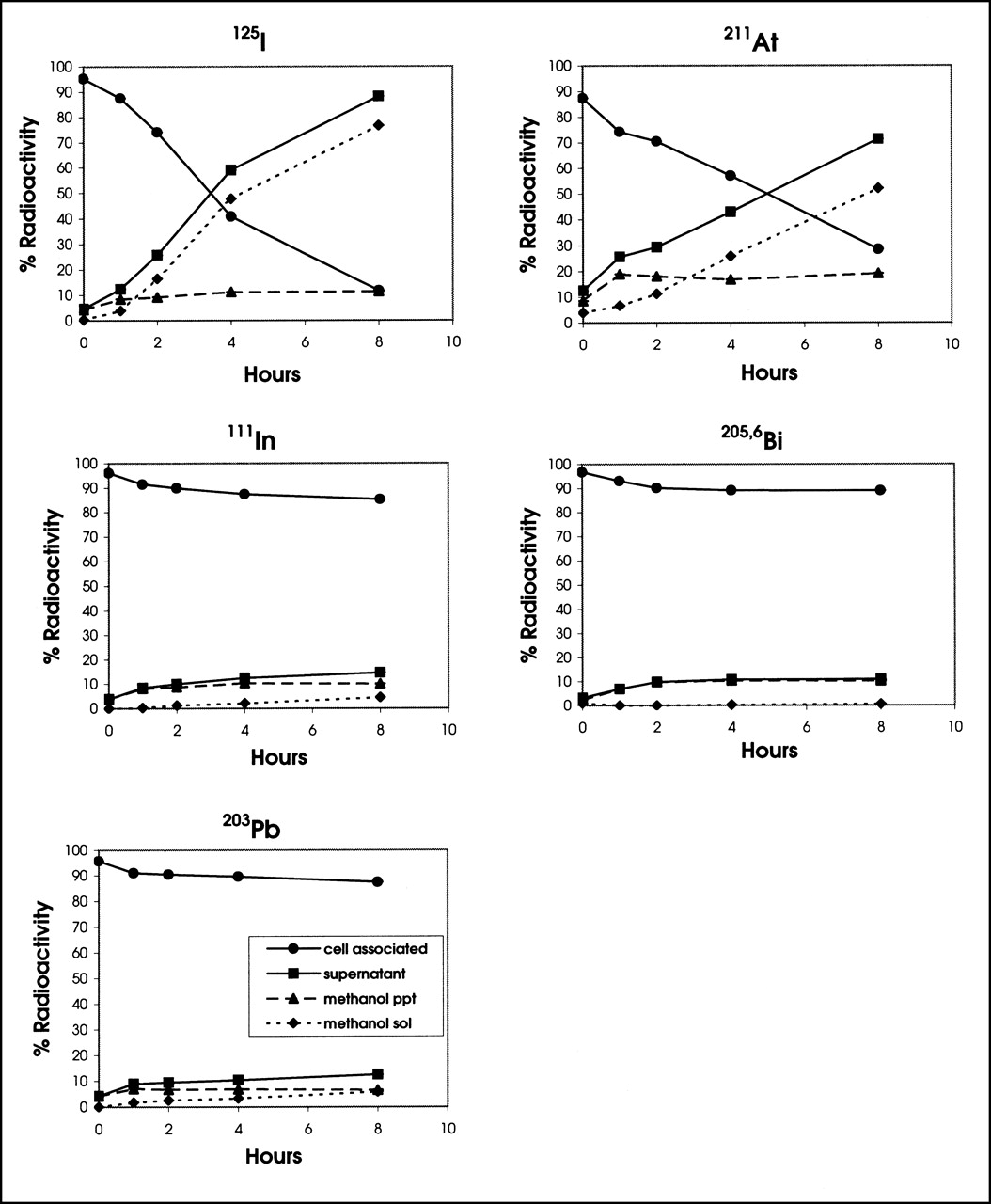
Although quantitatively different, the trends of MOLT-4 cell retention and release of radioactivity into the supernatant for all 5 radionuclides were similar to those of PBMNC (Fig. 2). The retention of 111In, 203Pb, and 205,6Bi by MOLT-4 malignant cells was high and in the range seen for PBMNC. The retention of 125I and 211At decreased when incubated at 37°C (Fig. 2). Accordingly, the amount of activity of both 125I and 211At increased in the supernatant of MOLT-4 cells, predominantly in the form of non–protein-bound radioactivity.
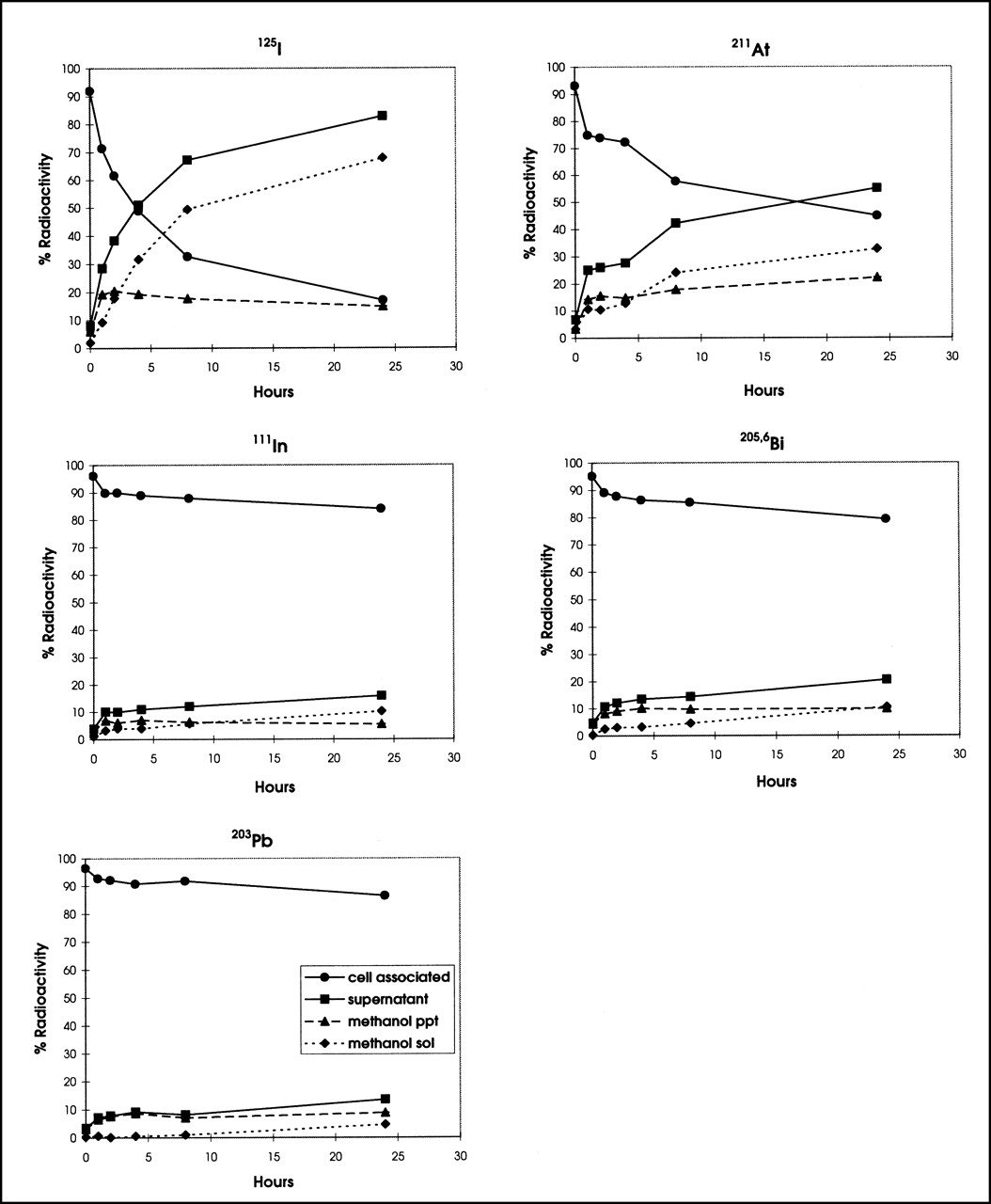
Retention and release of various γ- and α-emitter–labeled T101s by MOLT-4 cell line were determined as described for Figure 1.
The acid-resistant fraction represents the percentage of radioactivity that is internalized and retained inside the cell after the radiolabeled T101 is allowed to bind to the cell surface. The results with PBMNC are shown in Figure 3. Before incubation at 37°C, the acid-resistant fractions were similar among the different radiolabels and most of the cell-associated radioactivity was on the cell surface and, therefore, removable with the acidic solution. One hour after incubation, the acid-resistant fraction increased. For 111In, 203Pb, and 205,6Bi, the amount continued to increase and started to level off at 2 h, reflecting the rapid internalization of the radiolabeled T101 and the cell retention of these radionuclides. In contrast, 125I and 211At showed a decrease in cellular retention after 2 h, reflecting the internalization, antibody catabolism, and subsequent release of the radionuclides.
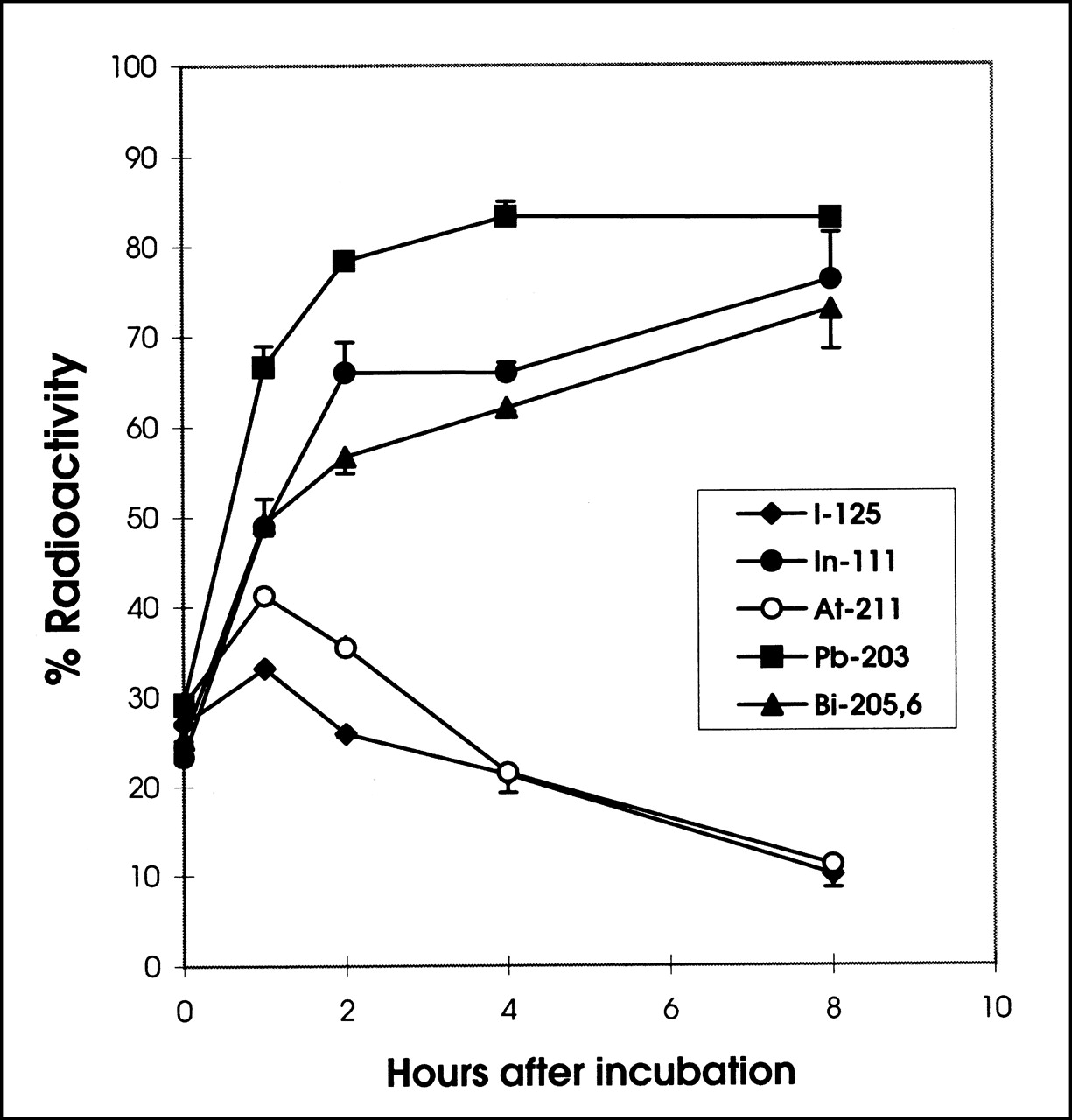
Percentage (mean ± SD) of acid-resistant to total cell-bound radioactivity for radiolabeled T101 by PBMNC. Labeled T101 was allowed to bind to PBMNC at 4°C for 1 h to obtain surface labeling with minimal internalization. Aliquots of 106 cells were then placed into separate tubes and supernatants were separated from cells. Immediately after separation, cells underwent acid wash and acid-resistant radioactivity was determined by counting cell pellets after centrifugation. This procedure was repeated at various times after transferring cell aliquots to incubator at 37°C. Acid-resistant fraction was determined by dividing activity in cell pellet after acid wash by total activity on cell after 1 h of incubation at 4°C.
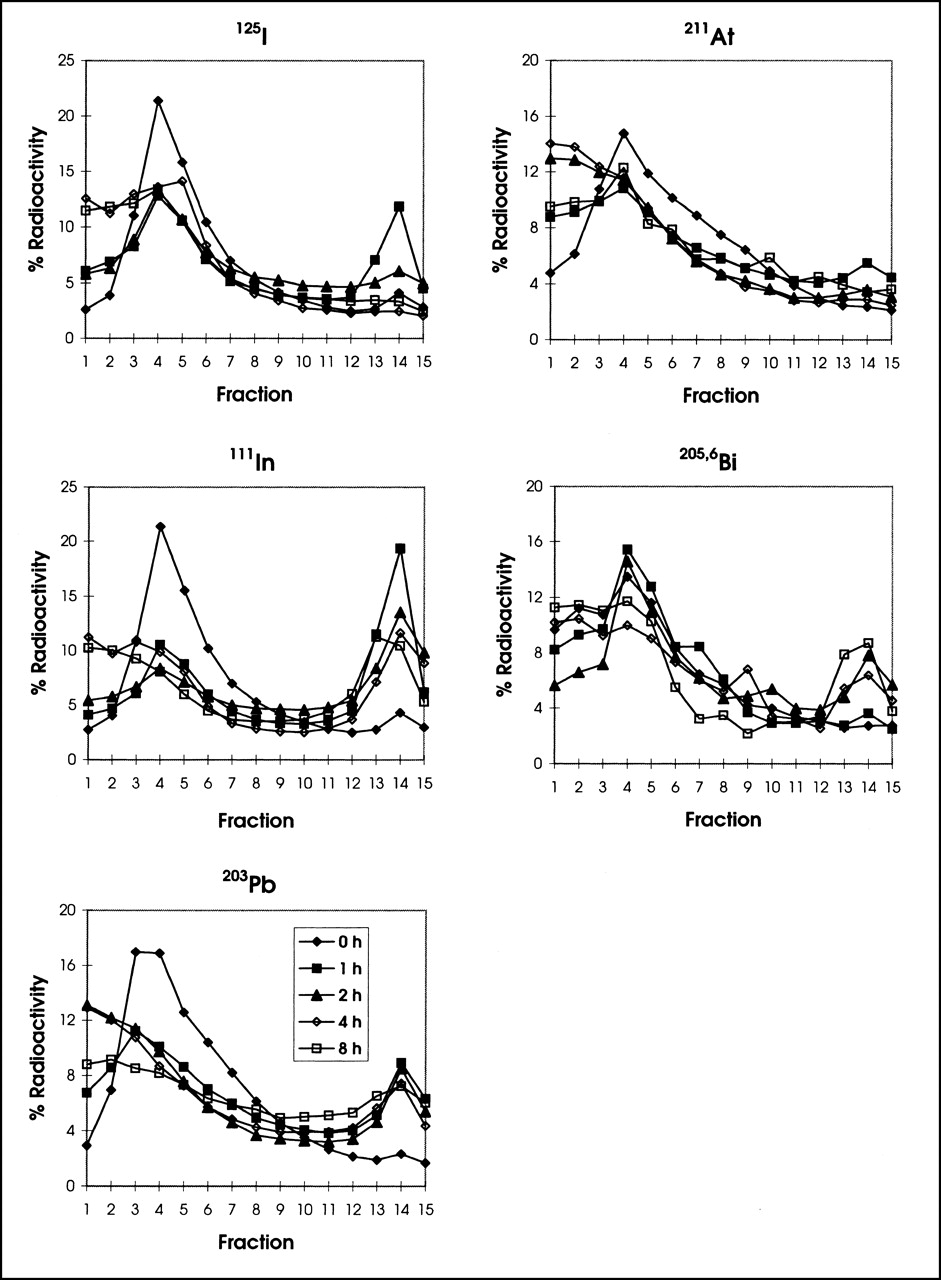
The cellular distribution of the various radionuclides was determined by Percoll gradient centrifugation after the cells were disrupted (Fig. 4). The radioactivity in the high-density lysosomal fraction (No. 14) increased 1 h after incubation for all preparations. The radioactivity in this fraction remained high throughout the 8-h period examined for the 111In, 203Pb, and 205,6Bi radiolabels, but decreased with 125I and 211At.

Intracellular distribution of 125I-, 211At-, 111In-, 205,6Bi-, and 203Pb-labeled T101 in PBMNC. PBMNC incubated with radiolabeled T101 were suspended in TES buffer and disrupted. Cell nuclei and unbroken cells were removed by centrifugation at 250g. Supernatants were applied to Percoll/TES buffer and ultracentrifuged at 4°C for 60 min at 20,000g. Serial 0.5 mL fractions were collected from top and counted for radioactivity. Early fractions represent activity on cell surface and late fractions represent activity in lysosomes (fraction 14). Location of lysosome fraction was confirmed using enzyme marker β-galactosidase (data not shown).
Discussion
The variable retention of radionuclides by target and nontarget cells has been recognized to be dependent on the radionuclides (6,7,31). In particular, with a rapidly internalizing antibody, cell retention of radionuclides is an important issue and may make the difference between an effective and ineffective antibody-based therapy (7). In addition, even for slowly internalizing antibodies, the retention of radioactivity varies depending on the radionuclide. With the developing interest in the use of α-emitting radionuclides for therapy (32–34), understanding their fate is important once internalization occurs because release from the target cells could result in unwanted localization and toxicity and in less dose being delivered to the target cells. The fate of the most promising α-emitting radionuclides, 2 of which are currently in clinical trials, was studied (32,33). Their catabolism was compared with that of 111In- and 125I-labeled T101, which are excellent prototypes of radiolabeled antibodies that have been evaluated both in vitro and in vivo (6,7). These 111In- and 125I-mAbs have shown large differences in radionuclide retention by targeted cells and therefore were used as a baseline comparison of the 2 extremes. Because of the short half-life or availability of 212Bi, 213Bi, and 212Pb, 205,6Bi and 203Pb were chosen as surrogate markers for these respective radionuclides.
This study showed rapid internalization of radiolabeled T101, as evidenced by the high cell-associated radioactivity over time of the radiometal isotopes (Figs. 1–3) and the lack of radioactivity released when an acid wash was used to remove cell surface radioactivity (Fig. 3). Further evidence of internalization was the high percentage of 125I and 211At present in the supernatants derived from catabolized antibody (Figs. 1 and 2). We used an acid wash buffer that was relatively mild (29) compared with that used by others (35) to minimize cell lysis. It is possible that this milder buffer resulted in less cell lysis and also less release from the cell surface than would have resulted from the use of other, harsher buffers.
Previous studies have shown that iodine from directly iodinated antibodies was rapidly released from cells when the antibody was catabolized, whereas radiometals such as 111In, 90Y, and 177Lu were retained once internalized. As expected, the chelated 205,6Bi and 203Pb behaved similarly to 111In. In PBMNC, the 205,6Bi and 203Pb radiometals showed little release from the cells. This pattern was similar to that observed with MOLT-4 cells. Some differences in quantitative rates of internalization and catabolism were expected because differences in internalization rates for PBMNC and malignant cells have been described with T101 (16,36) and with other antibodies (37).
The release of 211At from the cells (Figs. 1 and 2) was significantly faster than that of 205,6Bi, particularly in the PBMNC (Fig. 1). The activity released appeared in the supernatant predominantly in the form of non–protein-bound activity, indicating catabolism. This pattern was similar to that of 125I-T101, although 211At is retained in slightly greater amounts than 125I. These results indicate that, similar to 125I-T101, 211At-T101 undergoes catabolism after internalization and the radionuclides are rapidly released from the cells. Previous studies evaluating the internalization and catabolism of 211At-labeled anti–epidermal growth factor receptor mAb and the iodinated counterpart have also shown evidence of internalization and release of catabolites into the supernatant, although in that case the release of 211At was greater than that of 125I (30). This contrasts with the radiometals in which those radionuclides are retained for a prolonged period within the cells.
Our subcellular localization studies showed that, over time, the radionuclides migrated into the lysosomal fraction, where progressive accumulation of the radiometals was observed. In contrast, for 125I and 211At, selective accumulation does not occur in the lysosomes because of the rapid release of the 125I and 211At during catabolism of the radiolabeled mAb. Although several reports show routing to lysosomes with retention of radiometals in them, others have reported accumulation of 99mTc in the cytoplasm (38). Because the main purpose of this study was to determine the retention of the different radionuclides within targeted cells, detailed analysis to determine the chemical nature of the catabolites was not performed.
These studies suggest that, for internalizing antibodies, CHX-A″-DPTA or -DOTA conjugates used for bismuth or lead isotopes, respectively, are adequate and would result in long retention of the tracer in the target cells. Although this may not be as important for short-lived radionuclides like 213Bi, it may be significant for longer-lived radionuclides like 211At and 212Pb. Although 203Pb showed prolonged intracellular retention, it is theoretically possible that when 212Pb decays to its 212Bi daughter, the 212Bi is released from the chelate. Whether 212Bi released from the chelate would be retained intracellularly was not evaluated in this study. In contrast, as with 125I, the 211At labeling method is suboptimal with internalizing antibodies and would result in rapid release of the isotope from the target cells if internalized (7). This study indicates that alternative labeling strategies for 211At should be pursued. This would be analogous to the pursuit of dilactitol-tyramine labeling for iodinated antibodies that resulted in much longer cellular retention (39,40).
Conclusion
The 211At-antibody labeled using SAPS conjugation behaved similarly to the 125I-labeled antibody, with rapid release of the label from targeted cells after internalization, whereas radiometal α-emitters showed prolonged retention within target cells and thus were more likely to deliver a larger therapeutic dose to the target tissue. These findings, together with other previous reports, suggest that, as a group, similar radiometals attached to mAbs through an appropriately stable chelating agent will be retained intracellularly.
Acknowledgments
The authors thank Dr. Martin W. Brechbiel for providing advice and expertise regarding the α-emitters and for comments on the article, Dr. Meili Zhang for technical assistance, discussion, and comments on the article, and Paul Plascjak for production of astatine and bismuth.
Footnotes
Received Jan. 25, 2001; revision accepted Jun. 13, 2001.
For correspondence and reprints contact: Jorge A. Carrasquillo, MD, Bldg. 10, Room 1C-496, 10 Center Dr., MSC 1180, Bethesda, MD 20892-1180.
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Optimized Methods for the Production of High-Purity 203Pb Using Electroplated Thallium Targets
- Noninvasive in vivo imaging of CD4 cells in simian-human immunodeficiency virus (SHIV)-infected nonhuman primates
- Radioimmunotherapy with {alpha}-Particle-Emitting 213Bi-C-Functionalized trans-Cyclohexyl-Diethylenetriaminepentaacetic Acid-Humanized 3S193 Is Enhanced by Combination with Paclitaxel Chemotherapy
- Renal Tubulointerstitial Changes after Internal Irradiation with {alpha}-Particle-Emitting Actinium Daughters
- Efforts to Control the Errant Products of a Targeted In vivo Generator
- The Promise of Targeted {alpha}-Particle Therapy
- Targeting of HER2 Antigen for the Treatment of Disseminated Peritoneal Disease
- Targeting Primary Human Ph+ B-Cell Precursor Leukemia-Engrafted SCID Mice Using Radiolabeled Anti-CD19 Monoclonal Antibodies
- Pretarget radiotherapy with an anti-CD25 antibody-streptavidin fusion protein was effective in therapy of leukemia/lymphoma xenografts
- Pretargeting radioimmunotherapy of a murine model of adult T-cell leukemia with the alpha -emitting radionuclide, bismuth 213